

Age-related epigenetic changes in hippocampal subregions of four animal models of Alzheimer's disease
- PMID: 29113959
- PMCID: PMC6863355
- DOI: 10.1016/j.mcn.2017.11.002
Age-related epigenetic changes in hippocampal subregions of four animal models of Alzheimer's disease
Abstract
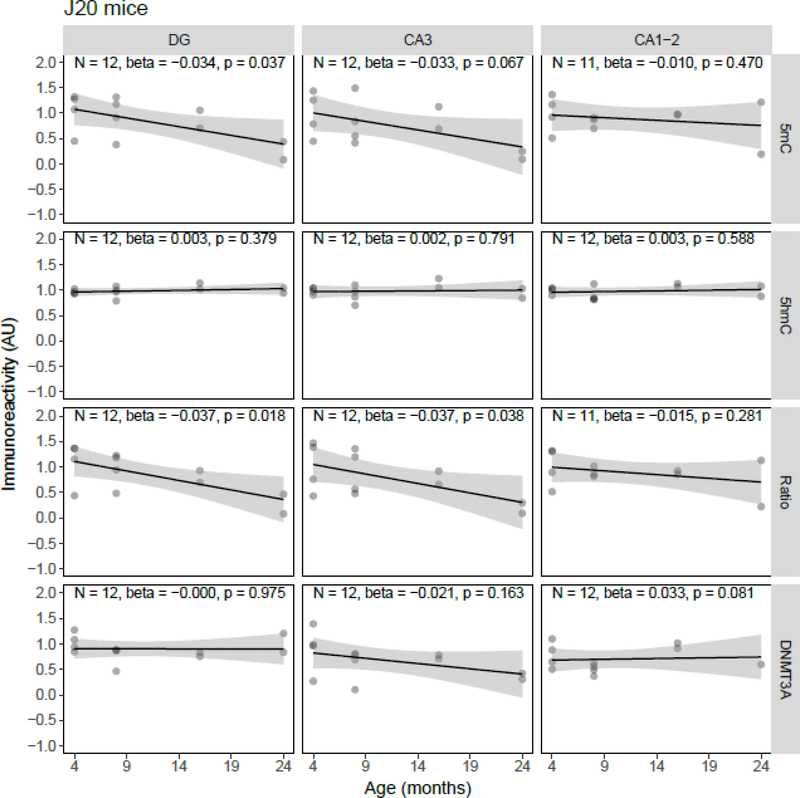
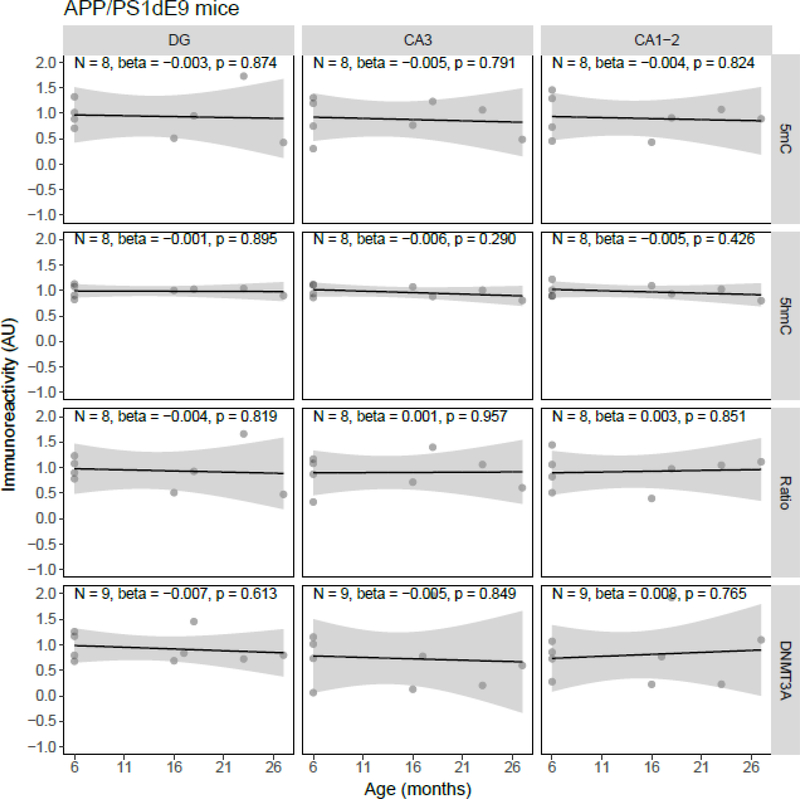
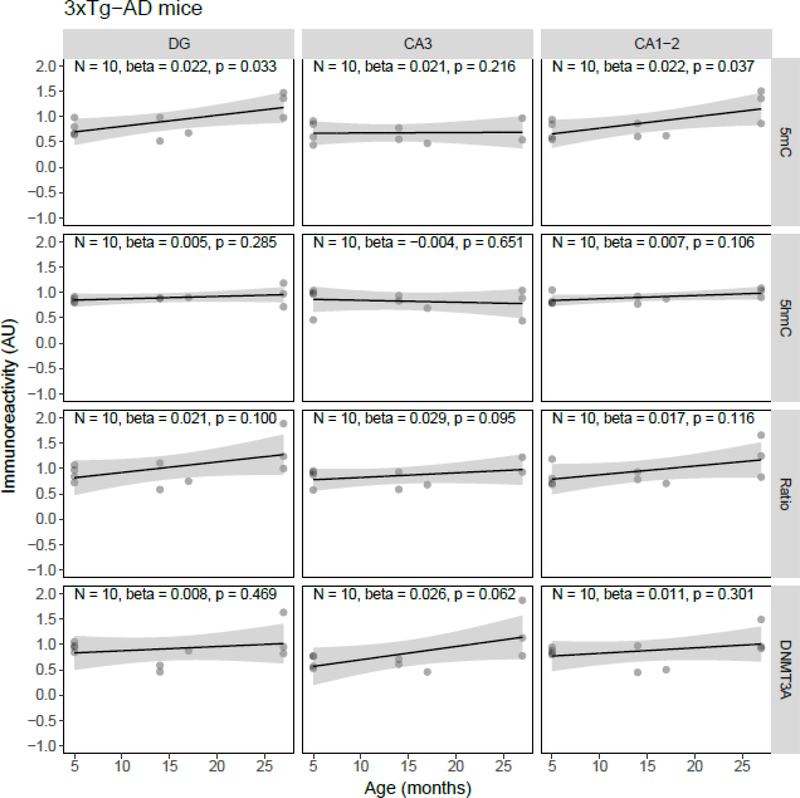
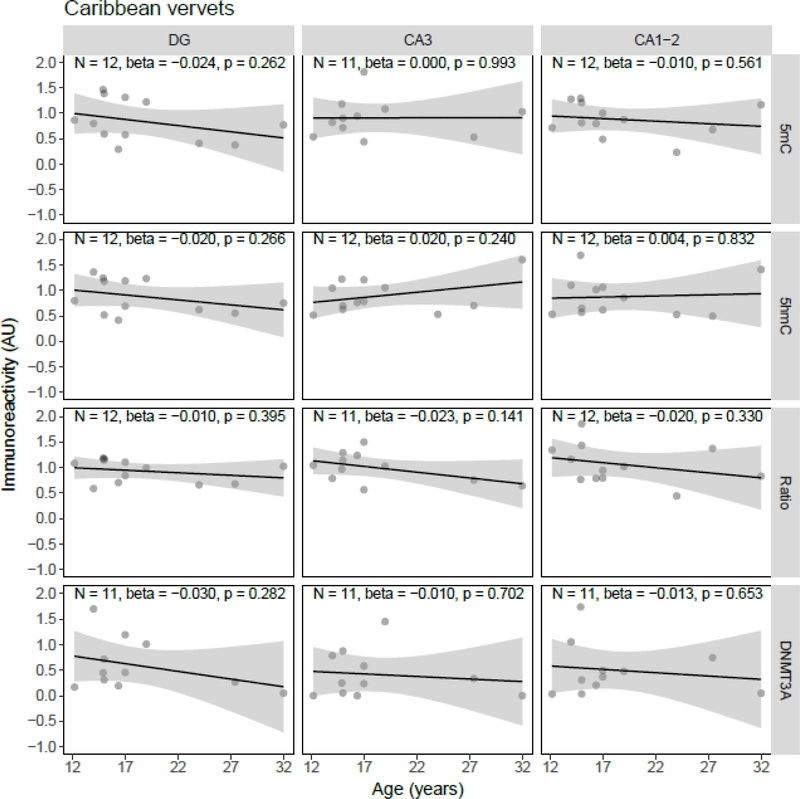
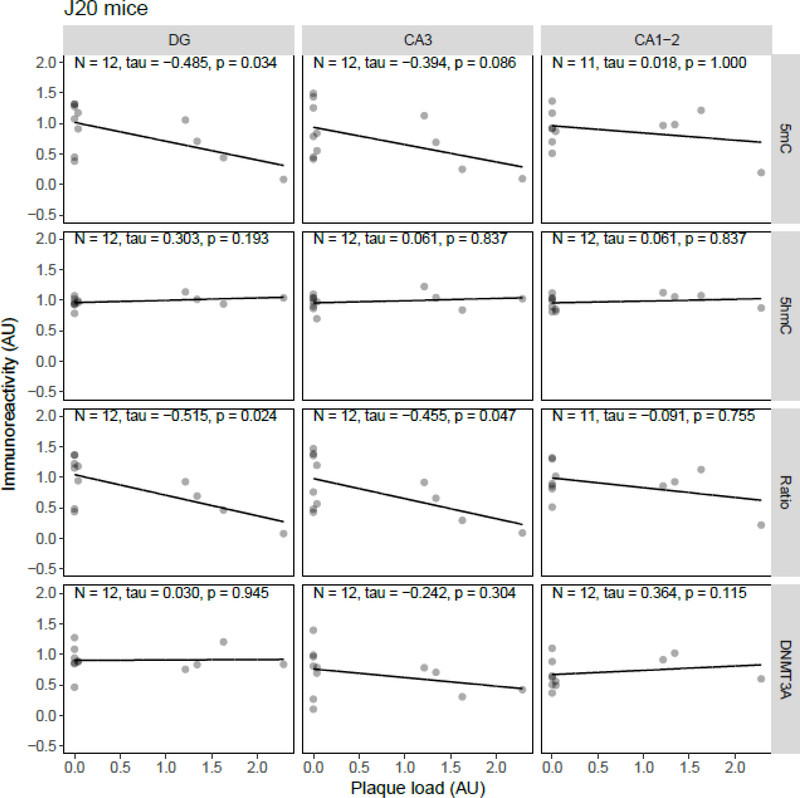
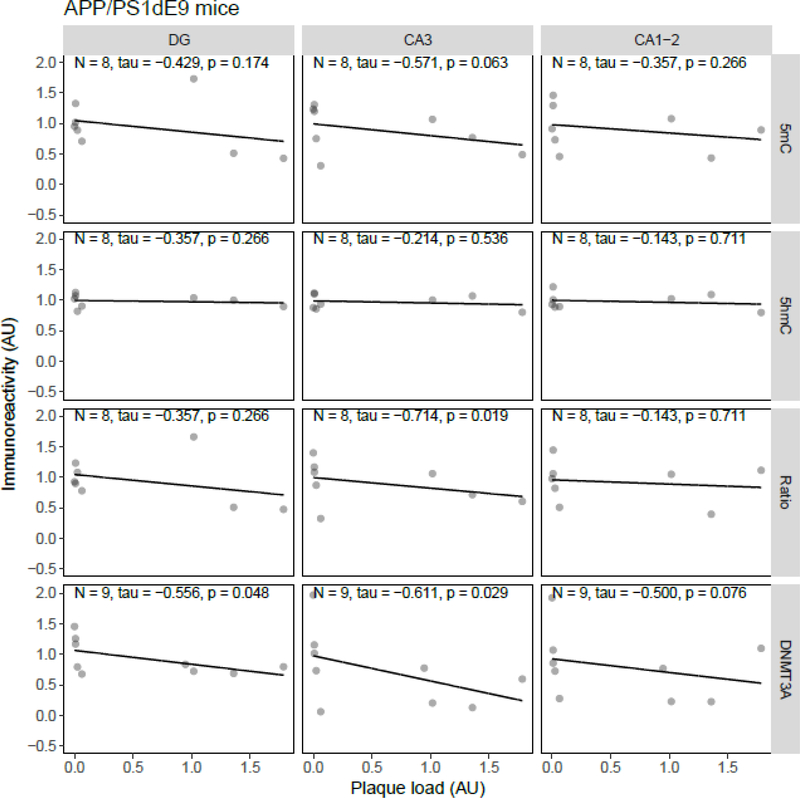
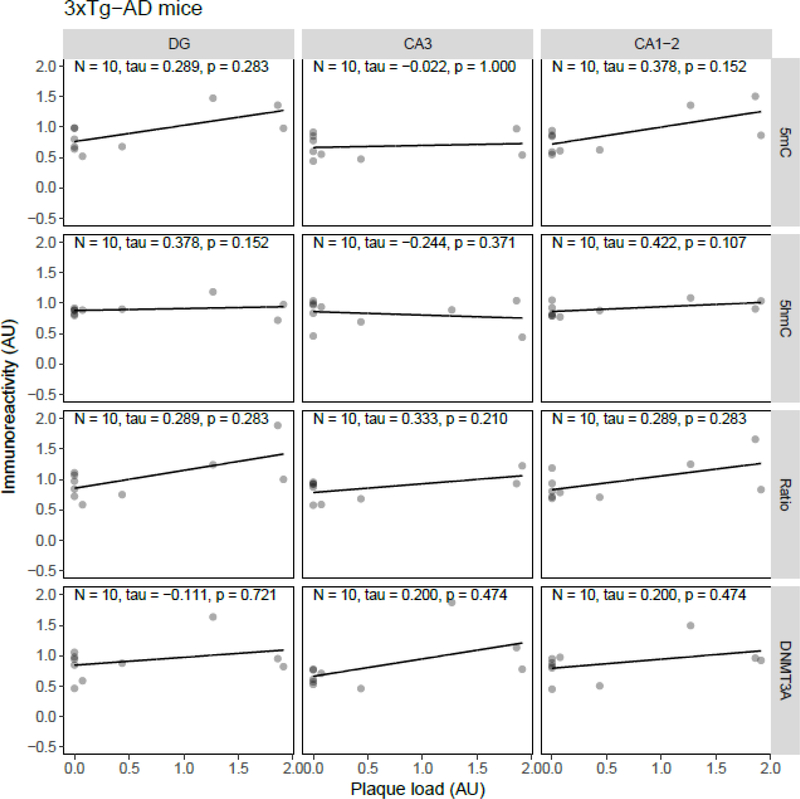
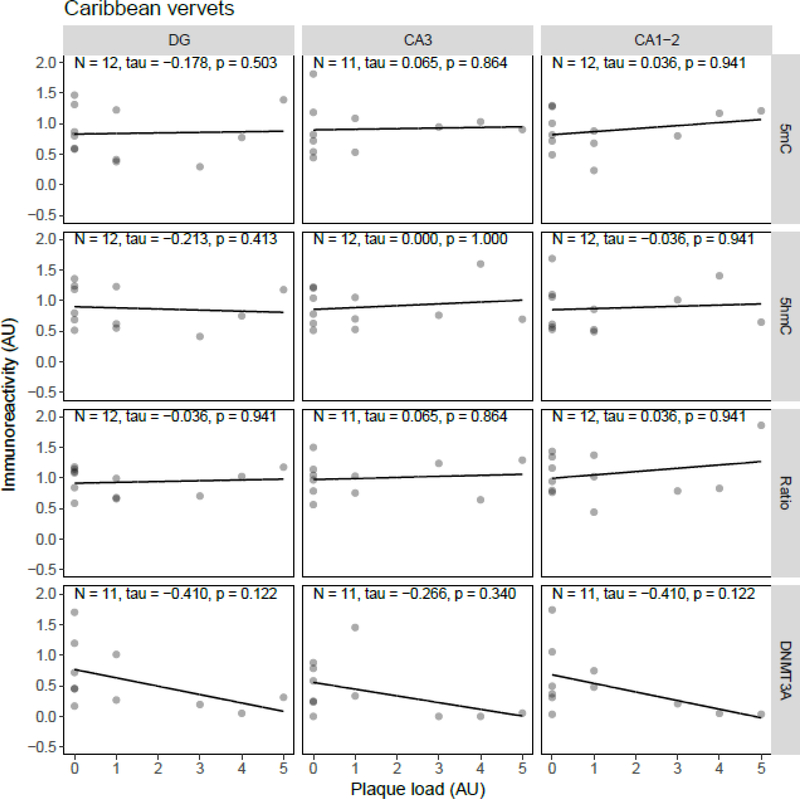
Both aging and Alzheimer's disease (AD) are associated with widespread epigenetic changes, with most evidence suggesting global hypomethylation in AD. It is, however, unclear how these age-related epigenetic changes are linked to molecular aberrations as expressed in animal models of AD. Here, we investigated age-related changes of epigenetic markers of DNA methylation and hydroxymethylation in a range of animal models of AD, and their correlations with amyloid plaque load. Three transgenic mouse models, including the J20, APP/PS1dE9 and 3xTg-AD models, as well as Caribbean vervets (a non-transgenic non-human primate model of AD) were investigated. In the J20 mouse model, an age-related decrease in DNA methylation was found in the dentate gyrus (DG) and a decrease in the ratio between DNA methylation and hydroxymethylation was found in the DG and cornu ammonis (CA) 3. In the 3xTg-AD mice, an age-related increase in DNA methylation was found in the DG and CA1-2. No significant age-related alterations were found in the APP/PS1dE9 mice and non-human primate model. In the J20 model, hippocampal plaque load showed a significant negative correlation with DNA methylation in the DG, and with the ratio a negative correlation in the DG and CA3. For the APP/PS1dE9 model a negative correlation between the ratio and plaque load was observed in the CA3, as well as a negative correlation between DNA methyltransferase 3A (DNMT3A) levels and plaque load in the DG and CA3. Thus, only the J20 model showed an age-related reduction in global DNA methylation, while DNA hypermethylation was observed in the 3xTg-AD model. Given these differences between animal models, future studies are needed to further elucidate the contribution of different AD-related genetic variation to age-related epigenetic changes.
Keywords: Aging; Alzheimer's disease; Animal models; DNA hydroxymethylation; DNA methylation; DNA methyltransferase; Hippocampus.
Copyright © 2017. Published by Elsevier Inc.
Figures
Similar articles
-
Active Amyloid-β Vaccination Results in Epigenetic Changes in the Hippocampus of an Alzheimer's Disease-Like Mouse Model.Curr Alzheimer Res. 2019;16(9):861-870. doi: 10.2174/1567205016666190827122009. Curr Alzheimer Res. 2019. PMID: 31453788
-
Consistent decrease in global DNA methylation and hydroxymethylation in the hippocampus of Alzheimer's disease patients.Neurobiol Aging. 2013 Sep;34(9):2091-9. doi: 10.1016/j.neurobiolaging.2013.02.021. Epub 2013 Apr 9. Neurobiol Aging. 2013. PMID: 23582657 Free PMC article.
-
Reversible epigenetic histone modifications and Bdnf expression in neurons with aging and from a mouse model of Alzheimer's disease.Age (Dordr). 2013 Jun;35(3):519-31. doi: 10.1007/s11357-011-9375-5. Epub 2012 Jan 12. Age (Dordr). 2013. PMID: 22237558 Free PMC article.
-
Age-dependent alterations of the hippocampal cell composition and proliferative potential in the hAβPPSwInd-J20 mouse.J Alzheimers Dis. 2014;41(4):1177-92. doi: 10.3233/JAD-132717. J Alzheimers Dis. 2014. PMID: 24787919
-
Abnormal Homocysteine Metabolism: An Insight of Alzheimer's Disease from DNA Methylation.Behav Neurol. 2020 Sep 8;2020:8438602. doi: 10.1155/2020/8438602. eCollection 2020. Behav Neurol. 2020. PMID: 32963633 Free PMC article. Review.
Cited by
-
Dietary supplements in neurological diseases and brain aging.J Prev Med Hyg. 2022 Oct 17;63(2 Suppl 3):E174-E188. doi: 10.15167/2421-4248/jpmh2022.63.2S3.2759. eCollection 2022 Jun. J Prev Med Hyg. 2022. PMID: 36479494 Free PMC article. Review.
-
DNA Methylation Signature of Aging: Potential Impact on the Pathogenesis of Parkinson's Disease.J Parkinsons Dis. 2023;13(2):145-164. doi: 10.3233/JPD-223517. J Parkinsons Dis. 2023. PMID: 36710687 Free PMC article. Review.
-
Age-related Disturbances in DNA (hydroxy)methylation in APP/PS1 Mice.Transl Neurosci. 2018 Dec 31;9:190-202. doi: 10.1515/tnsci-2018-0028. eCollection 2018. Transl Neurosci. 2018. PMID: 30746282 Free PMC article.
-
Biological aging processes underlying cognitive decline and neurodegenerative disease.J Clin Invest. 2022 May 16;132(10):e158453. doi: 10.1172/JCI158453. J Clin Invest. 2022. PMID: 35575089 Free PMC article. Review.
-
Epigenetic Factors in Late-Onset Alzheimer's Disease: MTHFR and CTH Gene Polymorphisms, Metabolic Transsulfuration and Methylation Pathways, and B Vitamins.Int J Mol Sci. 2019 Jan 14;20(2):319. doi: 10.3390/ijms20020319. Int J Mol Sci. 2019. PMID: 30646578 Free PMC article. Review.
References
-
- Bottiglieri T, Godfrey P, Flynn T, Carney MW, Toone BK, Reynolds EH, 1990. Cerebrospinal fluid S-adenosylmethionine in depression and dementia: effects of treatment with parenteral and oral S-adenosylmethionine. J. Neurol. Neurosurg. Psychiatry 53, 1096–1098. doi:10.1136/jnnp.53.12.1096 - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
