

Identification of residues within the African swine fever virus DP71L protein required for dephosphorylation of translation initiation factor eIF2α and inhibiting activation of pro-apoptotic CHOP
- PMID: 28189088
- PMCID: PMC5346070
- DOI: 10.1016/j.virol.2017.02.002
Identification of residues within the African swine fever virus DP71L protein required for dephosphorylation of translation initiation factor eIF2α and inhibiting activation of pro-apoptotic CHOP
Abstract
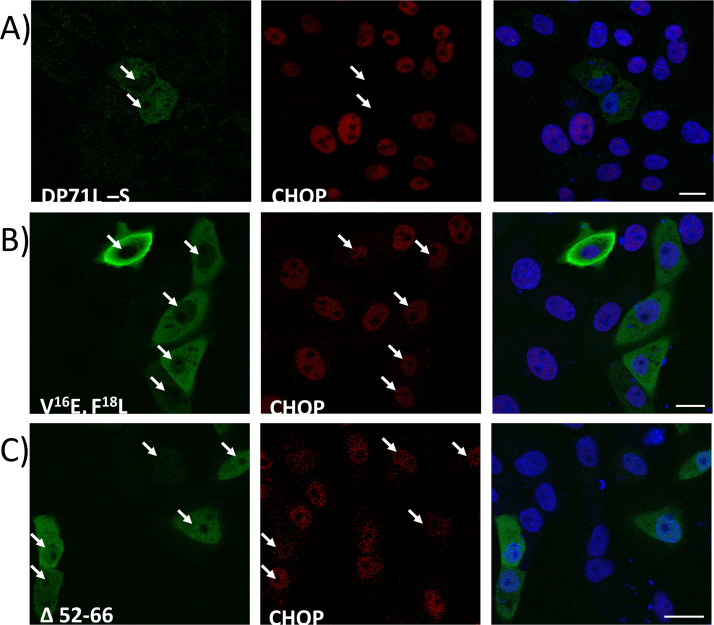
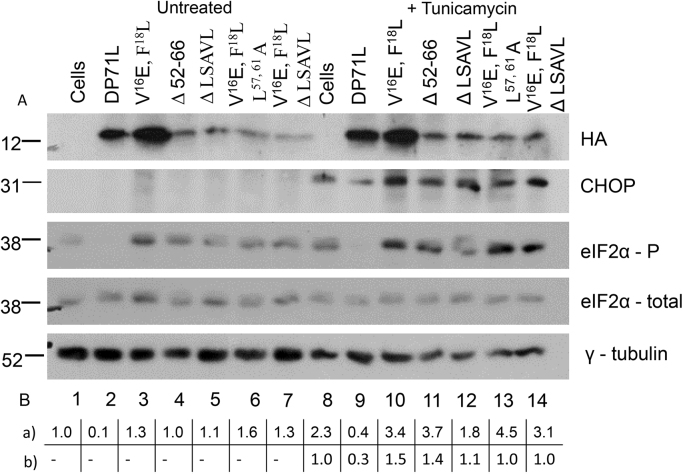
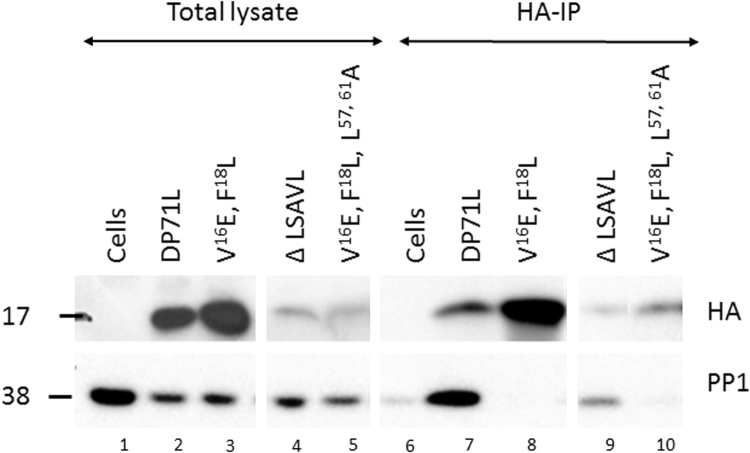
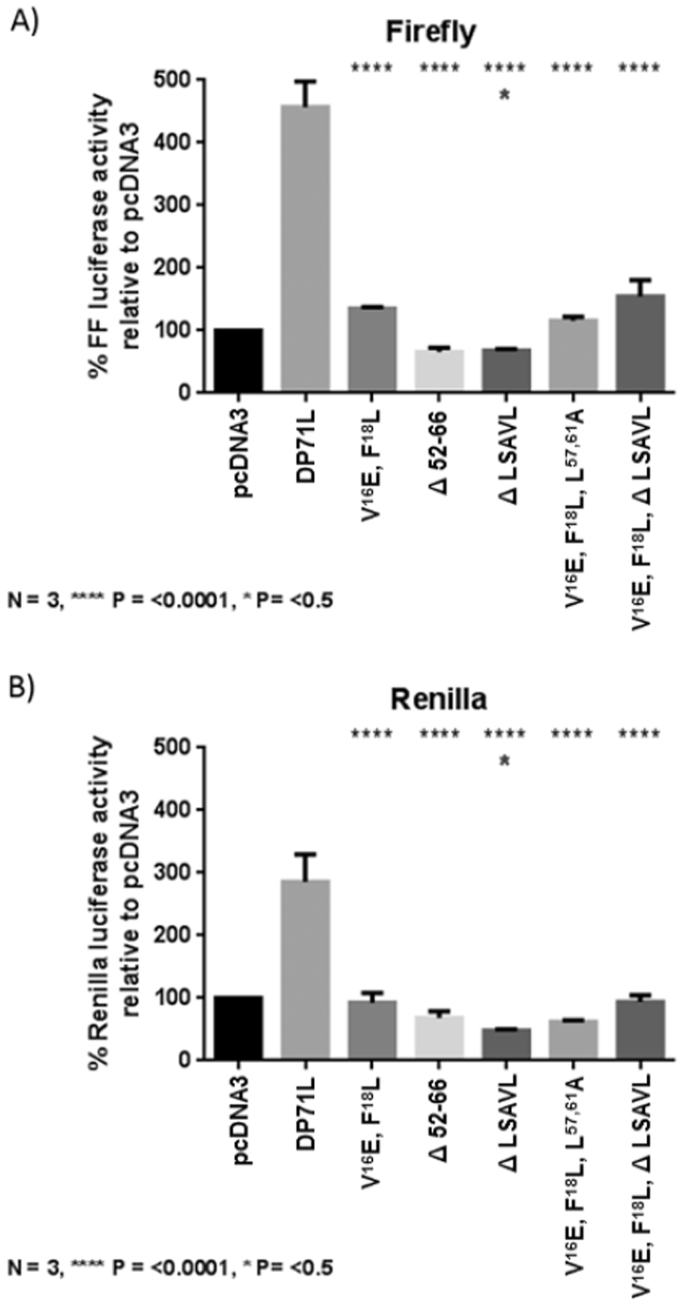
The African swine fever virus DP71L protein recruits protein phosphatase 1 (PP1) to dephosphorylate the translation initiation factor 2α (eIF2α) and avoid shut-off of global protein synthesis and downstream activation of the pro-apoptotic factor CHOP. Residues V16 and F18A were critical for binding of DP71L to PP1. Mutation of this PP1 binding motif or deletion of residues between 52 and 66 reduced the ability of DP71L to cause dephosphorylation of eIF2α and inhibit CHOP induction. The residues LSAVL, between 57 and 61, were also required. PP1 was co-precipitated with wild type DP71L and the mutant lacking residues 52- 66 or the LSAVL motif, but not with the PP1 binding motif mutant. The residues in the LSAVL motif play a critical role in DP71L function but do not interfere with binding to PP1. Instead we propose these residues are important for DP71L binding to eIF2α.
Keywords: African swine fever virus; CHOP; EIF2α; Protein translation; Unfolded protein response.
Copyright © 2017 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

Similar articles
-
The African swine fever virus DP71L protein recruits the protein phosphatase 1 catalytic subunit to dephosphorylate eIF2alpha and inhibits CHOP induction but is dispensable for these activities during virus infection.J Virol. 2010 Oct;84(20):10681-9. doi: 10.1128/JVI.01027-10. Epub 2010 Aug 11. J Virol. 2010. PMID: 20702639 Free PMC article.
-
The MyD116 African swine fever virus homologue interacts with the catalytic subunit of protein phosphatase 1 and activates its phosphatase activity.J Virol. 2007 Mar;81(6):2923-9. doi: 10.1128/JVI.02077-06. Epub 2007 Jan 10. J Virol. 2007. PMID: 17215279 Free PMC article.
-
An eIF2α-binding motif in protein phosphatase 1 subunit GADD34 and its viral orthologs is required to promote dephosphorylation of eIF2α.Proc Natl Acad Sci U S A. 2015 Jul 7;112(27):E3466-75. doi: 10.1073/pnas.1501557112. Epub 2015 Jun 22. Proc Natl Acad Sci U S A. 2015. PMID: 26100893 Free PMC article.
-
Investigations of Pro- and Anti-Apoptotic Factors Affecting African Swine Fever Virus Replication and Pathogenesis.Viruses. 2017 Aug 25;9(9):241. doi: 10.3390/v9090241. Viruses. 2017. PMID: 28841179 Free PMC article. Review.
-
An Overview of Methods for Detecting eIF2α Phosphorylation and the Integrated Stress Response.Methods Mol Biol. 2022;2428:3-18. doi: 10.1007/978-1-0716-1975-9_1. Methods Mol Biol. 2022. PMID: 35171470 Review.
Cited by
-
The A179L Gene of African Swine Fever Virus Suppresses Virus-Induced Apoptosis but Enhances Necroptosis.Viruses. 2021 Dec 13;13(12):2490. doi: 10.3390/v13122490. Viruses. 2021. PMID: 34960759 Free PMC article.
-
Nonstructural protein NS17 of grass carp reovirus Honghu strain promotes virus infection by mediating cell-cell fusion and apoptosis.Virus Res. 2023 Sep;334:199150. doi: 10.1016/j.virusres.2023.199150. Epub 2023 Jun 16. Virus Res. 2023. PMID: 37302658 Free PMC article.
-
The role of host eIF2α in viral infection.Virol J. 2020 Jul 23;17(1):112. doi: 10.1186/s12985-020-01362-6. Virol J. 2020. PMID: 32703221 Free PMC article. Review.
-
Infection of domestic pigs with a genotype II potent strain of ASFV causes cytokine storm and lymphocyte mass reduction.Front Immunol. 2024 Apr 18;15:1361531. doi: 10.3389/fimmu.2024.1361531. eCollection 2024. Front Immunol. 2024. PMID: 38698849 Free PMC article.
-
Emerging roles of the Protein Phosphatase 1 (PP1) in the context of viral infections.Cell Commun Signal. 2024 Jan 24;22(1):65. doi: 10.1186/s12964-023-01468-8. Cell Commun Signal. 2024. PMID: 38267954 Free PMC article. Review.
References
-
- Brush M.H., Weiser D.C., Shenolikar S. Growth arrest and DNA damage-inducible protein GADD34 targets protein phosphatase 1 alpha to the endoplasmic reticulum and promotes dephosphorylation of the alpha subunit of eukaryotic translation initiation factor 2. Mol. Cell. Biol. 2003;23:1292–1303. - PMC - PubMed
MeSH terms
Substances
Grants and funding
- BB/E019781/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/E021239/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00001709/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BBS/E/I/00007037/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
