

Tumor Suppressor Interferon-Regulatory Factor 1 Counteracts the Germinal Center Reaction Driven by a Cancer-Associated Gammaherpesvirus
- PMID: 26719266
- PMCID: PMC4810652
- DOI: 10.1128/JVI.02774-15
Tumor Suppressor Interferon-Regulatory Factor 1 Counteracts the Germinal Center Reaction Driven by a Cancer-Associated Gammaherpesvirus
Abstract
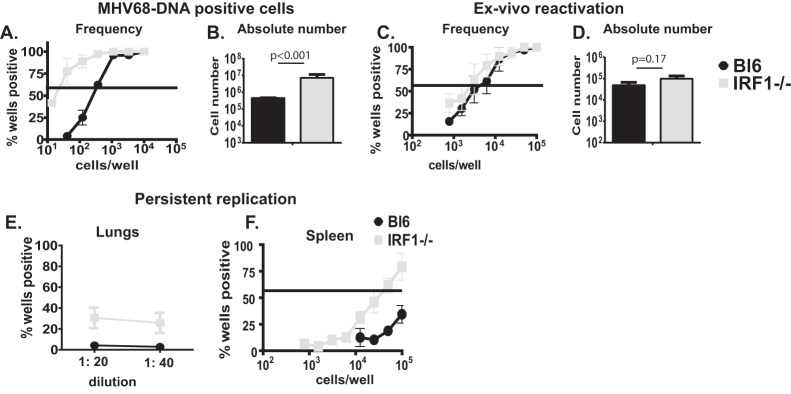
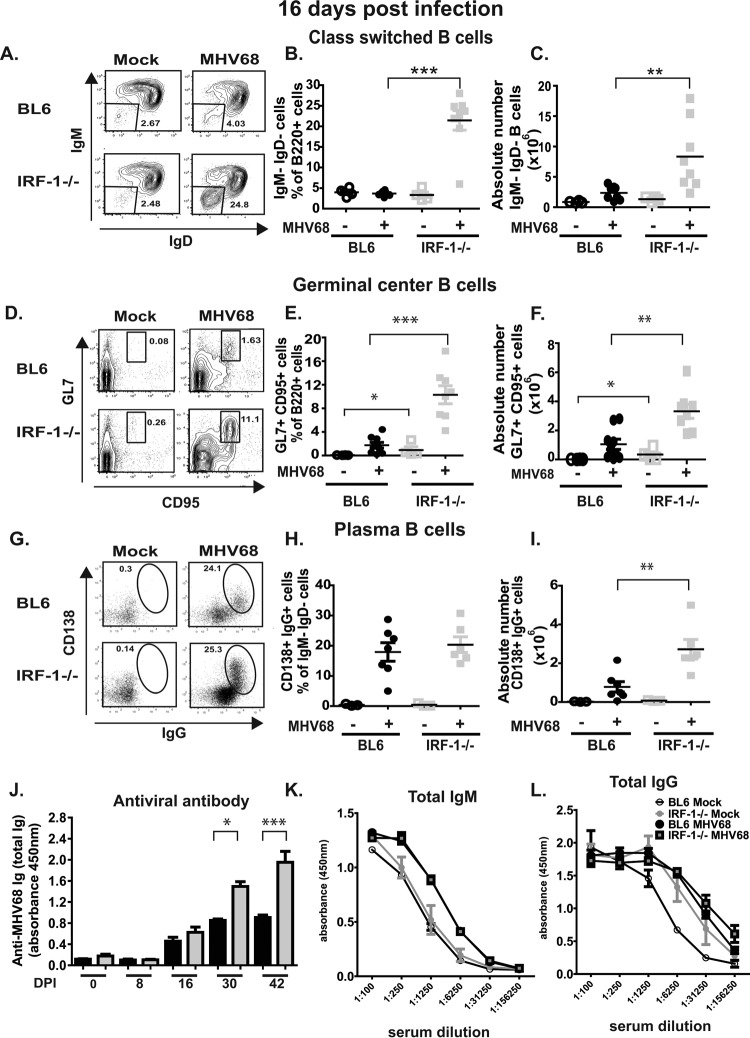
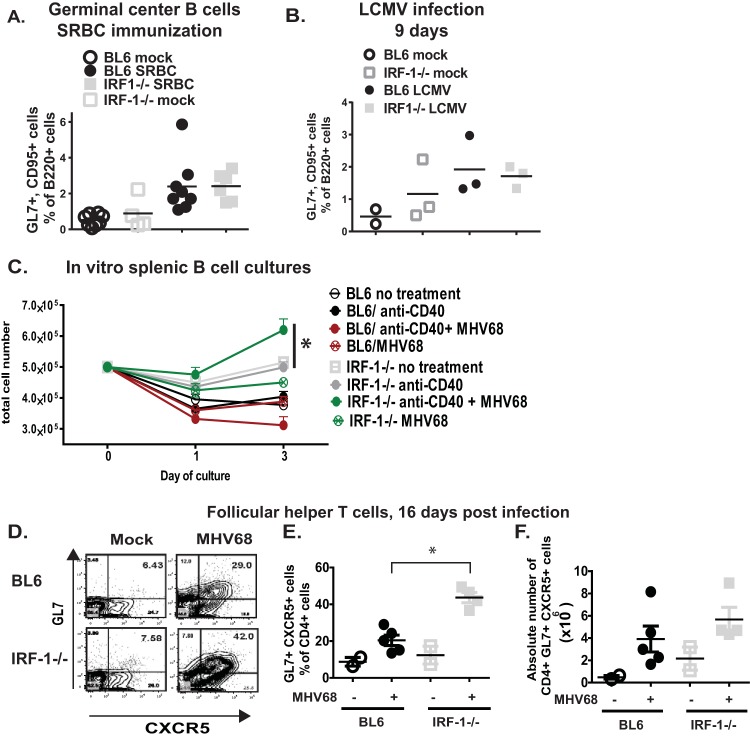
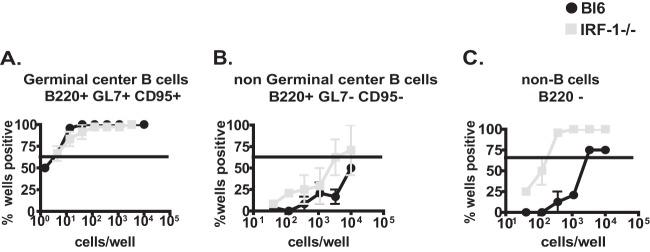
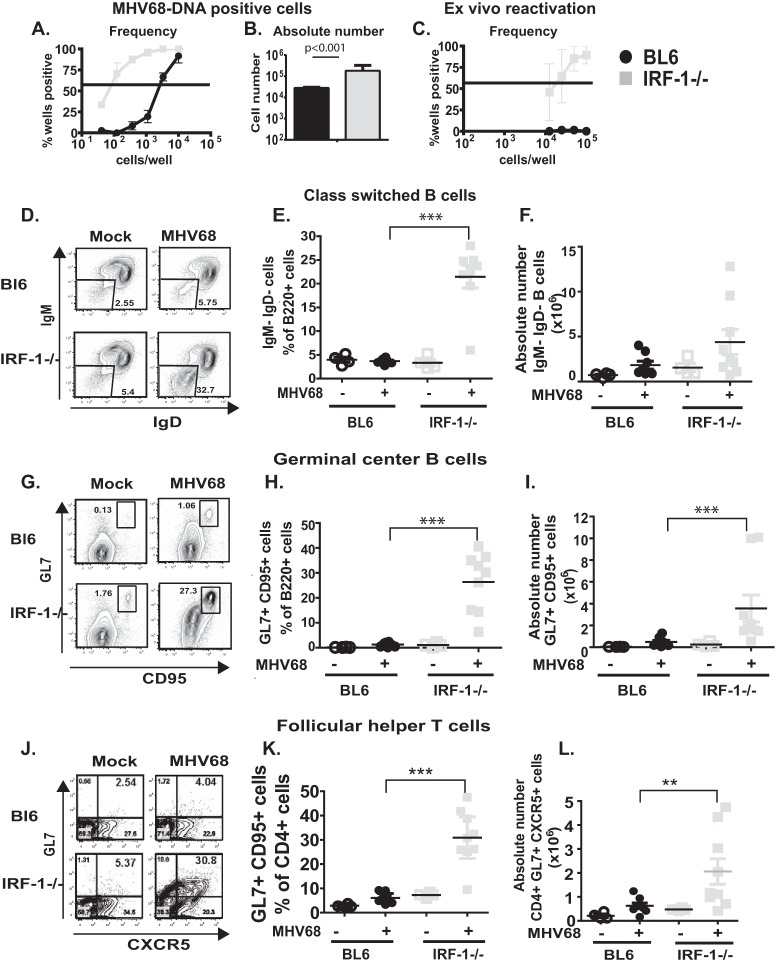
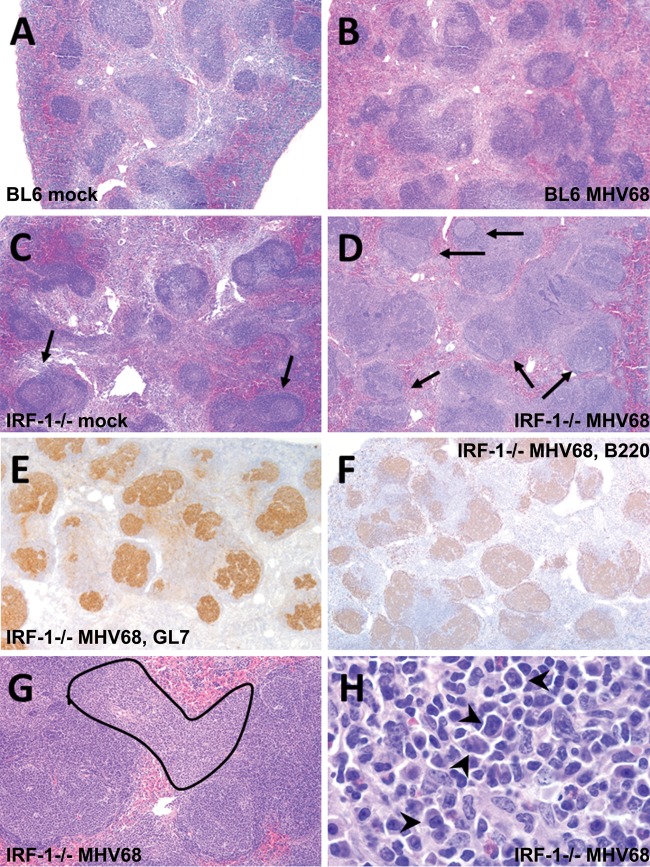
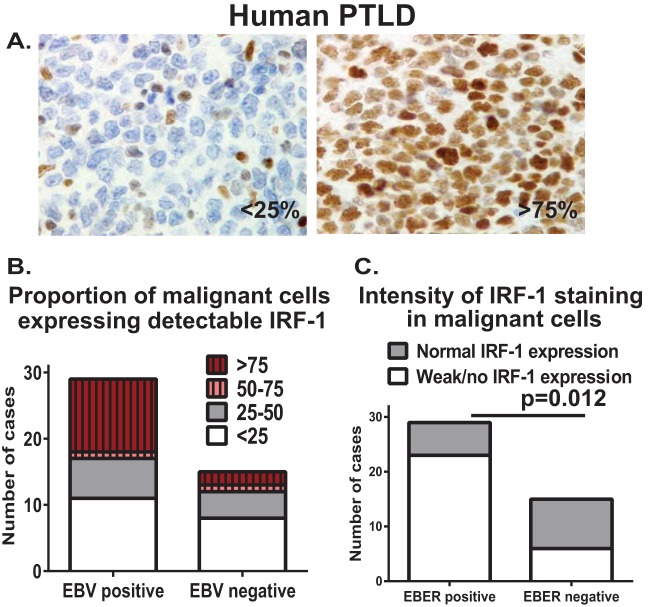
Gammaherpesviruses are ubiquitous pathogens that are associated with the development of B cell lymphomas. Gammaherpesviruses employ multiple mechanisms to transiently stimulate a broad, polyclonal germinal center reaction, an inherently mutagenic stage of B cell differentiation that is thought to be the primary target of malignant transformation in virus-driven lymphomagenesis. We found that this gammaherpesvirus-driven germinal center expansion was exaggerated and lost its transient nature in the absence of interferon-regulatory factor 1 (IRF-1), a transcription factor with antiviral and tumor suppressor functions. Uncontrolled and persistent expansion of germinal center B cells led to pathological changes in the spleens of chronically infected IRF-1-deficient animals. Additionally, we found decreased IRF-1 expression in cases of human posttransplant lymphoproliferative disorder, a malignant condition associated with gammaherpesvirus infection. The results of our study define an unappreciated role for IRF-1 in B cell biology and provide insight into the potential mechanism of gammaherpesvirus-driven lymphomagenesis.
Importance: Gammaherpesviruses establish lifelong infection in most adults and are associated with B cell lymphomas. While the infection is asymptomatic in many hosts, it is critical to identify individuals who may be at an increased risk of virus-induced cancer. Such identification is currently impossible, as the host risk factors that predispose individuals toward viral lymphomagenesis are poorly understood. The current study identifies interferon-regulatory factor 1 (IRF-1) to be one of such candidate host factors. Specifically, we found that IRF-1 enforces long-term suppression of an inherently mutagenic stage of B cell differentiation that gammaherpesviruses are thought to target for transformation. Correspondingly, in the absence of IRF-1, chronic gammaherpesvirus infection induced pathological changes in the spleens of infected animals. Further, we found decreased IRF-1 expression in human gammaherpesvirus-induced B cell malignancies.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures
Similar articles
-
B Cell-Intrinsic Expression of Interferon Regulatory Factor 1 Supports Chronic Murine Gammaherpesvirus 68 Infection.J Virol. 2020 Jun 16;94(13):e00399-20. doi: 10.1128/JVI.00399-20. Print 2020 Jun 16. J Virol. 2020. PMID: 32321819 Free PMC article.
-
T Cell-Intrinsic Interferon Regulatory Factor 1 Expression Suppresses Differentiation of CD4+ T Cell Populations That Support Chronic Gammaherpesvirus Infection.J Virol. 2021 Sep 27;95(20):e0072621. doi: 10.1128/JVI.00726-21. Epub 2021 Aug 4. J Virol. 2021. PMID: 34346769 Free PMC article.
-
Interferon Regulatory Factor 3 Supports the Establishment of Chronic Gammaherpesvirus Infection in a Route- and Dose-Dependent Manner.J Virol. 2021 Apr 12;95(9):e02208-20. doi: 10.1128/JVI.02208-20. Print 2021 Apr 12. J Virol. 2021. PMID: 33597211 Free PMC article.
-
Gammaherpesviruses and B Cells: A Relationship That Lasts a Lifetime.Viral Immunol. 2020 May;33(4):316-326. doi: 10.1089/vim.2019.0126. Epub 2020 Jan 8. Viral Immunol. 2020. PMID: 31913773 Free PMC article. Review.
-
Dangerous Liaisons: Gammaherpesvirus Subversion of the Immunoglobulin Repertoire.Viruses. 2020 Jul 23;12(8):788. doi: 10.3390/v12080788. Viruses. 2020. PMID: 32717815 Free PMC article. Review.
Cited by
-
Kaposi sarcoma-associated herpesvirus: immunobiology, oncogenesis, and therapy.J Clin Invest. 2016 Sep 1;126(9):3165-75. doi: 10.1172/JCI84418. Epub 2016 Sep 1. J Clin Invest. 2016. PMID: 27584730 Free PMC article. Review.
-
T Cell-Intrinsic Interleukin 17 Receptor A Signaling Supports the Establishment of Chronic Murine Gammaherpesvirus 68 Infection.J Virol. 2022 Jul 27;96(14):e0063922. doi: 10.1128/jvi.00639-22. Epub 2022 Jun 27. J Virol. 2022. PMID: 35758659 Free PMC article.
-
Tumor suppressor Interferon Regulatory Factor 1 selectively blocks expression of endogenous retrovirus.Virology. 2019 Jan 2;526:52-60. doi: 10.1016/j.virol.2018.10.003. Epub 2018 Oct 17. Virology. 2019. PMID: 30342302 Free PMC article.
-
Conserved Gammaherpesvirus Protein Kinase Selectively Promotes Irrelevant B Cell Responses.J Virol. 2019 Apr 3;93(8):e01760-18. doi: 10.1128/JVI.01760-18. Print 2019 Apr 15. J Virol. 2019. PMID: 30728267 Free PMC article.
-
Interferon Regulatory Factor 1 and Type I Interferon Cooperate To Control Acute Gammaherpesvirus Infection.J Virol. 2016 Dec 16;91(1):e01444-16. doi: 10.1128/JVI.01444-16. Print 2017 Jan 1. J Virol. 2016. PMID: 27795415 Free PMC article.
References
-
- Schoggins JW, MacDuff DA, Imanaka N, Gainey MD, Shrestha B, Eitson JL, Mar KB, Richardson RB, Ratushny AV, Litvak V, Dabelic R, Manicassamy B, Aitchison JD, Aderem A, Elliott RM, Garcia-Sastre A, Racaniello V, Snijder EJ, Yokoyama WM, Diamond MS, Virgin HW, Rice CM. 2014. Pan-viral specificity of IFN-induced genes reveals new roles for cGAS in innate immunity. Nature 505:691–695. doi:10.1038/nature14555. - DOI - PMC - PubMed
-
- Nair S, Michaelsen-Preusse K, Finsterbusch K, Stegemann-Koniszewski S, Bruder D, Grashoff M, Korte M, Koster M, Kalinke U, Hauser H, Kroger A. 2014. Interferon regulatory factor-1 protects from fatal neurotropic infection with vesicular stomatitis virus by specific inhibition of viral replication in neurons. PLoS Pathog 10:e1003999. doi:10.1371/journal.ppat.1003999. - DOI - PMC - PubMed
-
- Maloney NS, Thackray LB, Goel G, Hwang S, Duan E, Vachharajani P, Xavier R, Virgin HW. 2012. Essential cell-autonomous role for interferon (IFN) regulatory factor 1 in IFN-gamma-mediated inhibition of norovirus replication in macrophages. J Virol 86:12655–12664. doi:10.1128/JVI.01564-12. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
