

The neonatal Fc receptor, FcRn, as a target for drug delivery and therapy
- PMID: 25703189
- PMCID: PMC4544678
- DOI: 10.1016/j.addr.2015.02.005
The neonatal Fc receptor, FcRn, as a target for drug delivery and therapy
Abstract
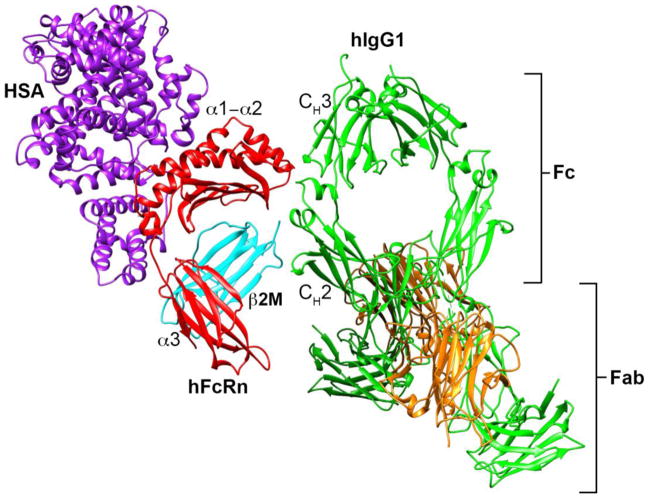
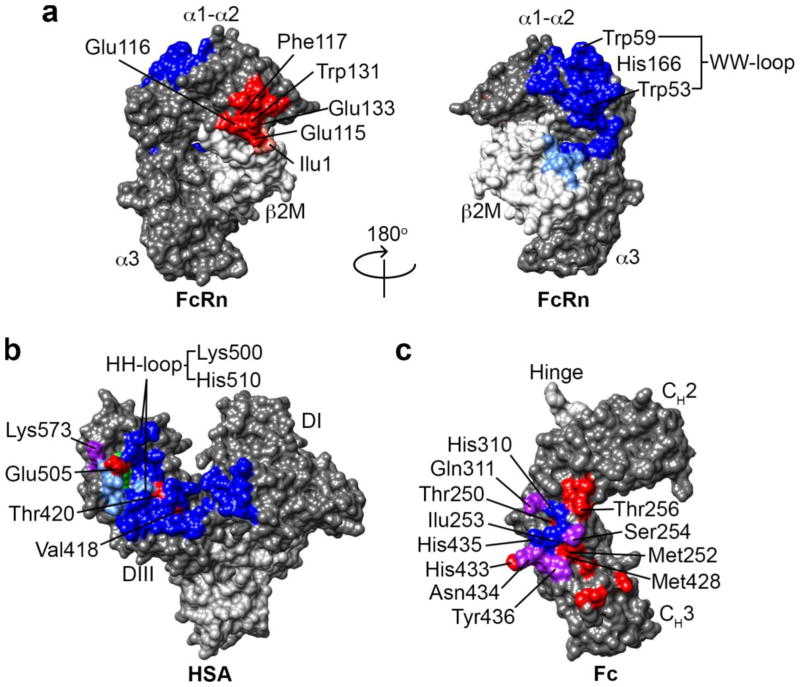
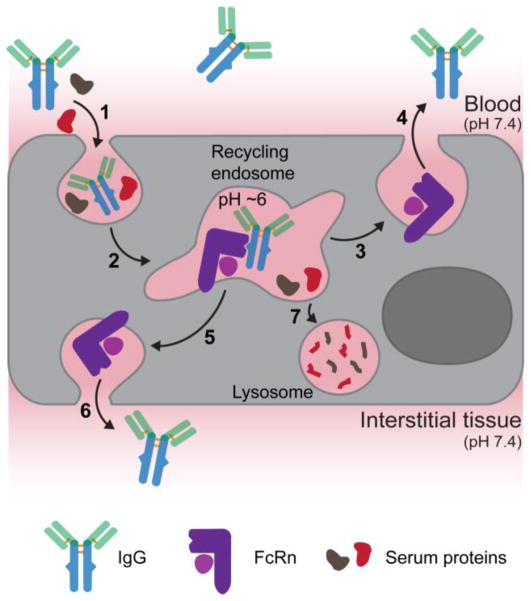
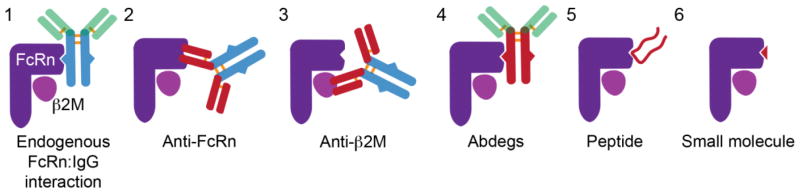
Immunoglobulin G (IgG)-based drugs are arguably the most successful class of protein therapeutics due in part to their remarkably long blood circulation. This arises from IgG interaction with the neonatal Fc receptor, FcRn. FcRn is the central regulator of IgG and albumin homeostasis throughout life and is increasingly being recognized as an important player in autoimmune disease, mucosal immunity, and tumor immune surveillance. Various engineering approaches that hijack or disrupt the FcRn-mediated transport pathway have been devised to develop long-lasting and non-invasive protein therapeutics, protein subunit vaccines, and therapeutics for treatment of autoimmune and infectious disease. In this review, we highlight the diverse biological functions of FcRn, emerging therapeutic opportunities, as well as the associated challenges of targeting FcRn for drug delivery and disease therapy.
Keywords: Albumin; FcRn; Immunoglobulin G; Nanoparticle; Protein engineering.
Copyright © 2015 Elsevier B.V. All rights reserved.
Conflict of interest statement
J.S. and F.C.S. declare no financial conflicts of interest.
Figures
Similar articles
-
Neonatal Fc receptor (FcRn): a novel target for therapeutic antibodies and antibody engineering.J Drug Target. 2014 May;22(4):269-78. doi: 10.3109/1061186X.2013.875030. Epub 2014 Jan 9. J Drug Target. 2014. PMID: 24404896 Review.
-
Qualification of a homogeneous cell-based neonatal Fc receptor (FcRn) binding assay and its application to studies on Fc functionality of IgG-based therapeutics.J Immunol Methods. 2013 Apr 30;390(1-2):81-91. doi: 10.1016/j.jim.2013.01.011. Epub 2013 Feb 4. J Immunol Methods. 2013. PMID: 23384837
-
The role of immunoglobulin transport receptor, neonatal Fc receptor in mucosal infection and immunity and therapeutic intervention.Int Immunopharmacol. 2024 Sep 10;138:112583. doi: 10.1016/j.intimp.2024.112583. Epub 2024 Jul 6. Int Immunopharmacol. 2024. PMID: 38971109 Review.
-
The Neonatal Fc Receptor (FcRn): A Misnomer?Front Immunol. 2019 Jul 10;10:1540. doi: 10.3389/fimmu.2019.01540. eCollection 2019. Front Immunol. 2019. PMID: 31354709 Free PMC article. Review.
-
Targeting FcRn for the modulation of antibody dynamics.Mol Immunol. 2015 Oct;67(2 Pt A):131-41. doi: 10.1016/j.molimm.2015.02.007. Epub 2015 Mar 9. Mol Immunol. 2015. PMID: 25766596 Free PMC article. Review.
Cited by
-
Charge heterogeneity: Basic antibody charge variants with increased binding to Fc receptors.MAbs. 2016 Nov/Dec;8(8):1548-1560. doi: 10.1080/19420862.2016.1225642. Epub 2016 Aug 25. MAbs. 2016. PMID: 27559765 Free PMC article.
-
Design of a Dual Agonist of Exendin-4 and FGF21 as a Potential Treatment for Type 2 Diabetes Mellitus and Obesity.Iran J Pharm Res. 2023 Aug 9;22(1):e131015. doi: 10.5812/ijpr-131015. eCollection 2023 Jan-Dec. Iran J Pharm Res. 2023. PMID: 38116563 Free PMC article.
-
Current Progress and Challenges in the Study of Adjuvants for Oral Vaccines.BioDrugs. 2023 Mar;37(2):143-180. doi: 10.1007/s40259-022-00575-1. Epub 2023 Jan 6. BioDrugs. 2023. PMID: 36607488 Free PMC article. Review.
-
Insertion of N-Terminal Hinge Glycosylation Enhances Interactions of the Fc Region of Human IgG1 Monomers with Glycan-Dependent Receptors and Blocks Hemagglutination by the Influenza Virus.J Immunol. 2019 Mar 1;202(5):1595-1611. doi: 10.4049/jimmunol.1801337. Epub 2019 Jan 25. J Immunol. 2019. PMID: 30683699 Free PMC article.
-
Engineering cytokines for cancer immunotherapy: a systematic review.Front Immunol. 2023 Jul 6;14:1218082. doi: 10.3389/fimmu.2023.1218082. eCollection 2023. Front Immunol. 2023. PMID: 37483629 Free PMC article. Review.
