

IgG subclasses and allotypes: from structure to effector functions
- PMID: 25368619
- PMCID: PMC4202688
- DOI: 10.3389/fimmu.2014.00520
IgG subclasses and allotypes: from structure to effector functions
Abstract
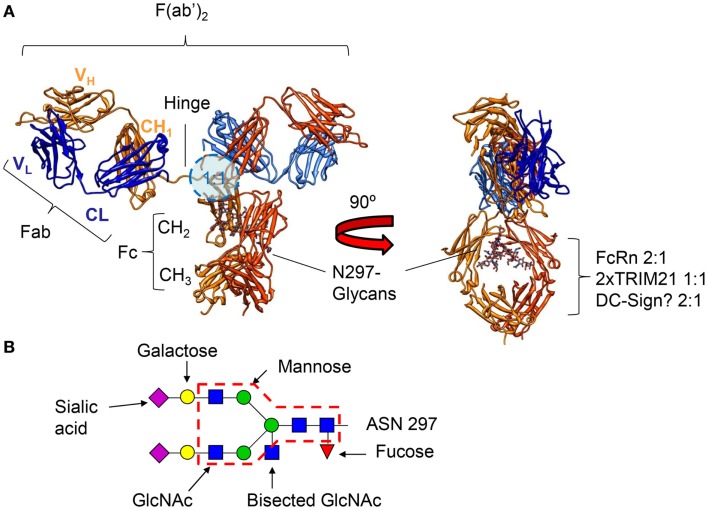
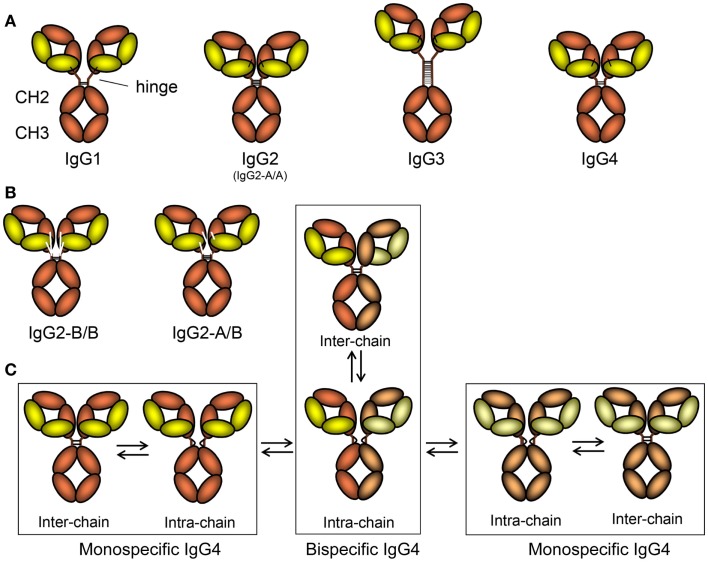
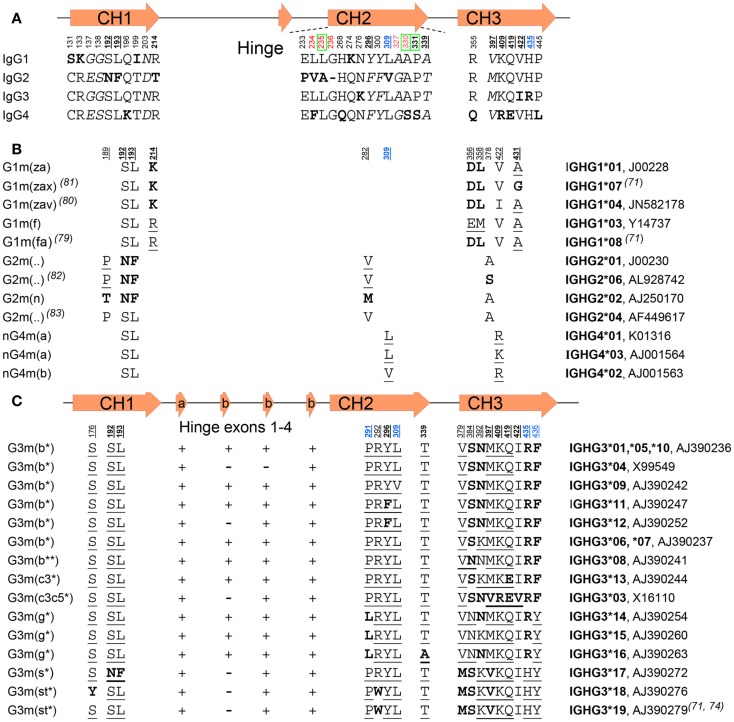
Of the five immunoglobulin isotypes, immunoglobulin G (IgG) is most abundant in human serum. The four subclasses, IgG1, IgG2, IgG3, and IgG4, which are highly conserved, differ in their constant region, particularly in their hinges and upper CH2 domains. These regions are involved in binding to both IgG-Fc receptors (FcγR) and C1q. As a result, the different subclasses have different effector functions, both in terms of triggering FcγR-expressing cells, resulting in phagocytosis or antibody-dependent cell-mediated cytotoxicity, and activating complement. The Fc-regions also contain a binding epitope for the neonatal Fc receptor (FcRn), responsible for the extended half-life, placental transport, and bidirectional transport of IgG to mucosal surfaces. However, FcRn is also expressed in myeloid cells, where it participates in both phagocytosis and antigen presentation together with classical FcγR and complement. How these properties, IgG-polymorphisms and post-translational modification of the antibodies in the form of glycosylation, affect IgG-function will be the focus of the current review.
Keywords: Fc receptors; IgG; genetic; glycosylation; immunoglobulin G; neonatal Fc receptor; polymorphism.
Figures
Similar articles
-
Fc-Galactosylation of Human Immunoglobulin Gamma Isotypes Improves C1q Binding and Enhances Complement-Dependent Cytotoxicity.Front Immunol. 2017 Jun 6;8:646. doi: 10.3389/fimmu.2017.00646. eCollection 2017. Front Immunol. 2017. PMID: 28634480 Free PMC article.
-
An engineered Fc variant of an IgG eliminates all immune effector functions via structural perturbations.Methods. 2014 Jan 1;65(1):114-26. doi: 10.1016/j.ymeth.2013.06.035. Epub 2013 Jul 17. Methods. 2014. PMID: 23872058
-
Structural characterization of the Man5 glycoform of human IgG3 Fc.Mol Immunol. 2017 Dec;92:28-37. doi: 10.1016/j.molimm.2017.10.001. Epub 2017 Oct 12. Mol Immunol. 2017. PMID: 29031045 Free PMC article.
-
How to select IgG subclasses in developing anti-tumor therapeutic antibodies.J Hematol Oncol. 2020 May 5;13(1):45. doi: 10.1186/s13045-020-00876-4. J Hematol Oncol. 2020. PMID: 32370812 Free PMC article. Review.
-
IgG-effector functions: "the good, the bad and the ugly".Immunol Lett. 2014 Aug;160(2):139-44. doi: 10.1016/j.imlet.2014.01.015. Epub 2014 Feb 1. Immunol Lett. 2014. PMID: 24495619 Review.
Cited by
-
Oropouche Virus Exposure in Febrile Patients during Chikungunya Virus Introduction in the State of Amapá, Amazon Region, Brazil.Pathogens. 2024 Jun 3;13(6):469. doi: 10.3390/pathogens13060469. Pathogens. 2024. PMID: 38921767 Free PMC article.
-
Selective Subnormal IgG1 in 54 Adult Index Patients with Frequent or Severe Bacterial Respiratory Tract Infections.J Immunol Res. 2016;2016:1405950. doi: 10.1155/2016/1405950. Epub 2016 Mar 31. J Immunol Res. 2016. PMID: 27123464 Free PMC article.
-
Bioorthogonal site-selective conjugation of fluorescent dyes to antibodies: method and potential applications.RSC Adv. 2022 Oct 5;12(44):28306-28317. doi: 10.1039/d2ra05580e. eCollection 2022 Oct 4. RSC Adv. 2022. PMID: 36320493 Free PMC article.
-
Maternal Immunization: Nature Meets Nurture.Front Microbiol. 2020 Jul 24;11:1499. doi: 10.3389/fmicb.2020.01499. eCollection 2020. Front Microbiol. 2020. PMID: 32849319 Free PMC article. Review.
-
In silico evaluation of the role of Fab glycosylation in cetuximab antibody dynamics.Front Immunol. 2024 Aug 8;15:1429600. doi: 10.3389/fimmu.2024.1429600. eCollection 2024. Front Immunol. 2024. PMID: 39185413 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials