

Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut
- PMID: 24799697
- PMCID: PMC4034235
- DOI: 10.1073/pnas.1323599111
Regulation of host weight gain and lipid metabolism by bacterial bile acid modification in the gut
Abstract
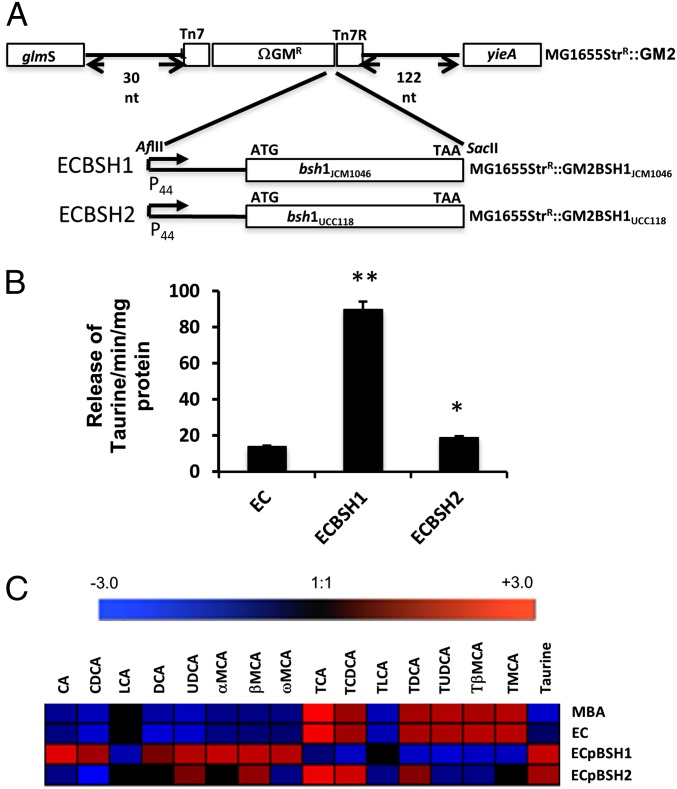
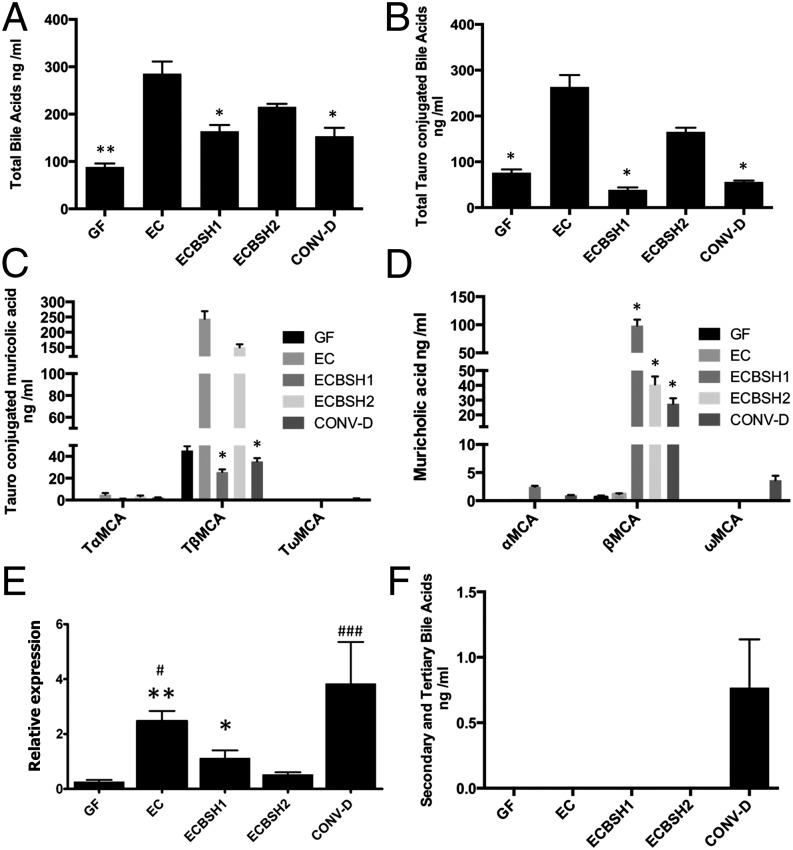
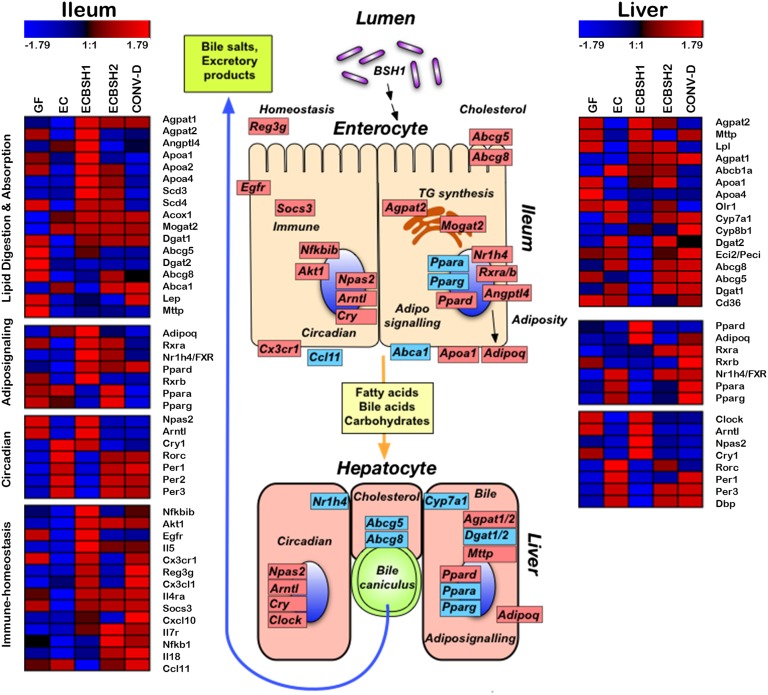
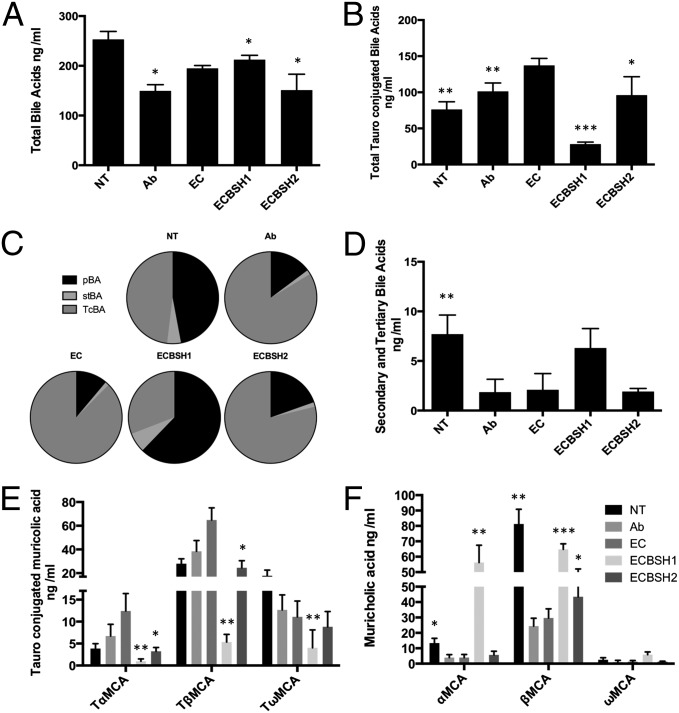
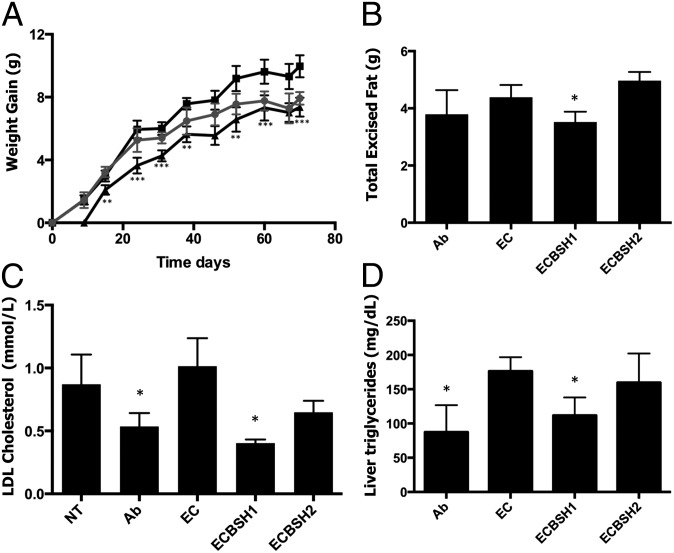
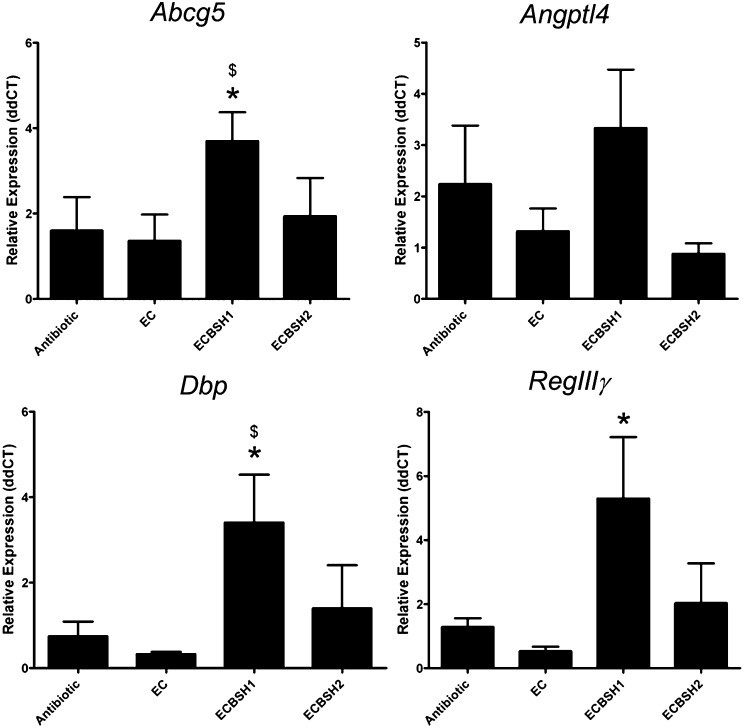
Alterations in the gastrointestinal microbiota have been implicated in obesity in mice and humans, but the key microbial functions influencing host energy metabolism and adiposity remain to be determined. Despite an increased understanding of the genetic content of the gastrointestinal microbiome, functional analyses of common microbial gene sets are required. We established a controlled expression system for the parallel functional analysis of microbial alleles in the murine gut. Using this approach we show that bacterial bile salt hydrolase (BSH) mediates a microbe-host dialogue that functionally regulates host lipid metabolism and plays a profound role in cholesterol metabolism and weight gain in the host. Expression of cloned BSH enzymes in the gastrointestinal tract of gnotobiotic or conventionally raised mice significantly altered plasma bile acid signatures and regulated transcription of key genes involved in lipid metabolism (Pparγ, Angptl4), cholesterol metabolism (Abcg5/8), gastrointestinal homeostasis (RegIIIγ), and circadian rhythm (Dbp, Per1/2) in the liver or small intestine. High-level expression of BSH in conventionally raised mice resulted in a significant reduction in host weight gain, plasma cholesterol, and liver triglycerides, demonstrating the overall impact of elevated BSH activity on host physiology. In addition, BSH activity in vivo varied according to BSH allele group, indicating that subtle differences in activity can have significant effects on the host. In summary, we demonstrate that bacterial BSH activity significantly impacts the systemic metabolic processes and adiposity in the host and represents a key mechanistic target for the control of obesity and hypercholesterolemia.
Keywords: FXR; Lactobacillus salivarius; adiponectin; barrier function; host response.
Conflict of interest statement
Conflict of interest statement: S.A.J., C.H., and C.G.M.G. have filed a patent relating to a portion of the work outlined in this paper.
Figures
Similar articles
-
Bacterial bile salt hydrolase in host metabolism: Potential for influencing gastrointestinal microbe-host crosstalk.Gut Microbes. 2014;5(5):669-74. doi: 10.4161/19490976.2014.969986. Gut Microbes. 2014. PMID: 25483337 Free PMC article. Review.
-
A selective gut bacterial bile salt hydrolase alters host metabolism.Elife. 2018 Jul 17;7:e37182. doi: 10.7554/eLife.37182. Elife. 2018. PMID: 30014852 Free PMC article.
-
Orally Administered Berberine Modulates Hepatic Lipid Metabolism by Altering Microbial Bile Acid Metabolism and the Intestinal FXR Signaling Pathway.Mol Pharmacol. 2017 Feb;91(2):110-122. doi: 10.1124/mol.116.106617. Epub 2016 Dec 8. Mol Pharmacol. 2017. PMID: 27932556 Free PMC article.
-
Bile Salt Hydrolase Activity-Based Probes for Monitoring Gut Microbial Bile Acid Metabolism.Chembiochem. 2024 May 17;25(10):e202300821. doi: 10.1002/cbic.202300821. Epub 2024 Apr 24. Chembiochem. 2024. PMID: 38564329
-
Interactions between gut bacteria and bile in health and disease.Mol Aspects Med. 2017 Aug;56:54-65. doi: 10.1016/j.mam.2017.06.002. Epub 2017 Jun 21. Mol Aspects Med. 2017. PMID: 28602676 Review.
Cited by
-
Interplay between Bile Acids and Intestinal Microbiota: Regulatory Mechanisms and Therapeutic Potential for Infections.Pathogens. 2024 Aug 20;13(8):702. doi: 10.3390/pathogens13080702. Pathogens. 2024. PMID: 39204302 Free PMC article. Review.
-
Multi-Omics Analysis Reveals Dietary Fiber's Impact on Growth, Slaughter Performance, and Gut Microbiome in Durco × Bamei Crossbred Pig.Microorganisms. 2024 Aug 14;12(8):1674. doi: 10.3390/microorganisms12081674. Microorganisms. 2024. PMID: 39203515 Free PMC article.
-
Effects of Synbiotic Administration on Gut Microbiome and Fecal Bile Acids in Dogs with Chronic Hepatobiliary Disease: A Randomized Case-Control Study.Vet Sci. 2024 Aug 10;11(8):364. doi: 10.3390/vetsci11080364. Vet Sci. 2024. PMID: 39195817 Free PMC article.
-
Navigating Challenges and Opportunities in Multi-Omics Integration for Personalized Healthcare.Biomedicines. 2024 Jul 5;12(7):1496. doi: 10.3390/biomedicines12071496. Biomedicines. 2024. PMID: 39062068 Free PMC article. Review.
-
Intersection of the microbiome and immune metabolism in lupus.Immunol Rev. 2024 Aug;325(1):77-89. doi: 10.1111/imr.13360. Epub 2024 Jun 14. Immunol Rev. 2024. PMID: 38873851 Review.
References
-
- Murphy EF, et al. Composition and energy harvesting capacity of the gut microbiota: Relationship to diet, obesity and time in mouse models. Gut. 2010;59(12):1635–1642. - PubMed
-
- Turnbaugh PJ, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–1031. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
