

Evidence that bank vole PrP is a universal acceptor for prions
- PMID: 24699458
- PMCID: PMC3974871
- DOI: 10.1371/journal.ppat.1003990
Evidence that bank vole PrP is a universal acceptor for prions
Abstract
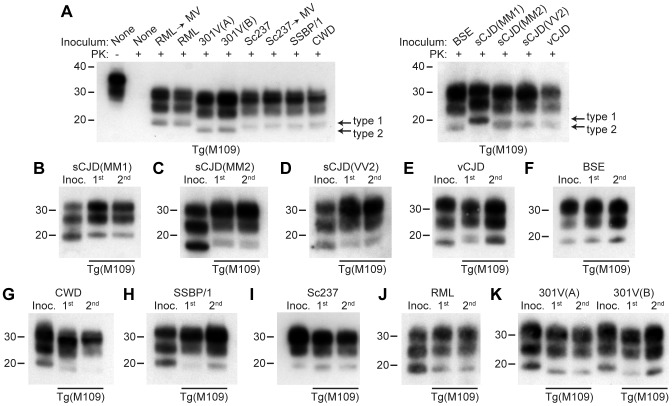
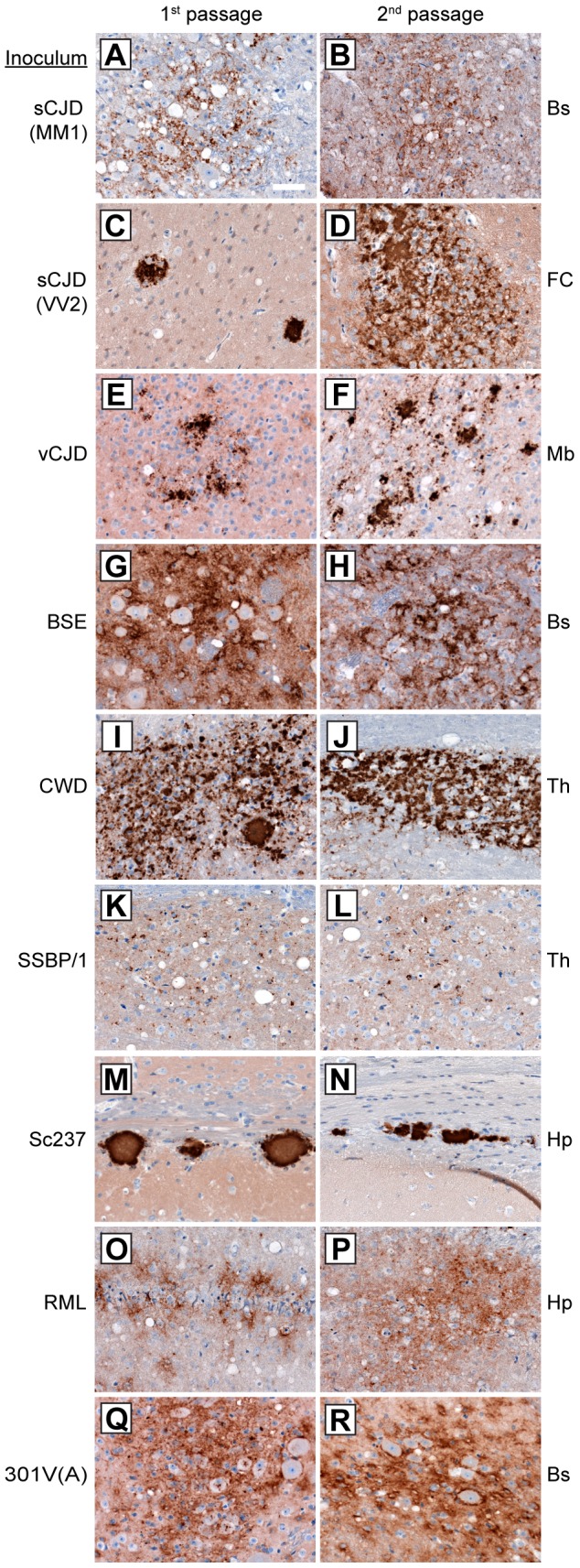
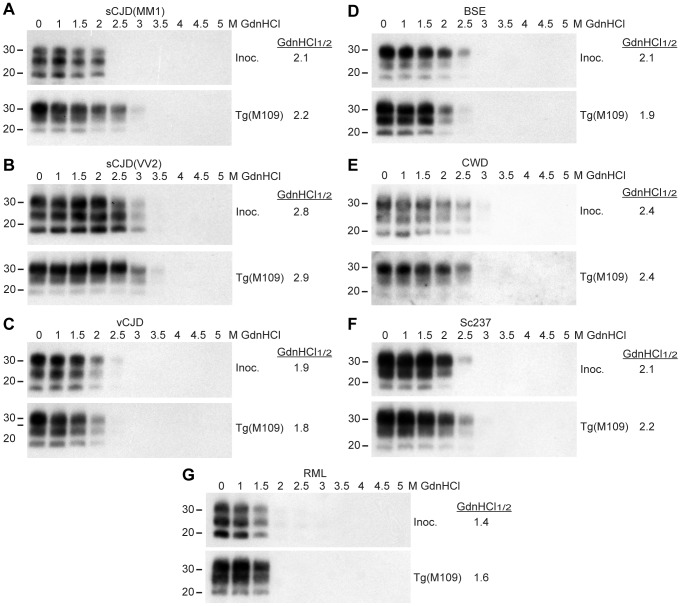
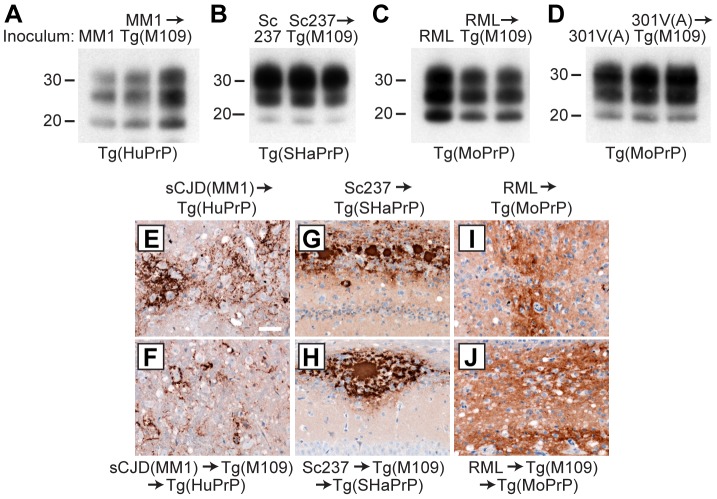
Bank voles are uniquely susceptible to a wide range of prion strains isolated from many different species. To determine if this enhanced susceptibility to interspecies prion transmission is encoded within the sequence of the bank vole prion protein (BVPrP), we inoculated Tg(M109) and Tg(I109) mice, which express BVPrP containing either methionine or isoleucine at polymorphic codon 109, with 16 prion isolates from 8 different species: humans, cattle, elk, sheep, guinea pigs, hamsters, mice, and meadow voles. Efficient disease transmission was observed in both Tg(M109) and Tg(I109) mice. For instance, inoculation of the most common human prion strain, sporadic Creutzfeldt-Jakob disease (sCJD) subtype MM1, into Tg(M109) mice gave incubation periods of ∼200 days that were shortened slightly on second passage. Chronic wasting disease prions exhibited an incubation time of ∼250 days, which shortened to ∼150 days upon second passage in Tg(M109) mice. Unexpectedly, bovine spongiform encephalopathy and variant CJD prions caused rapid neurological dysfunction in Tg(M109) mice upon second passage, with incubation periods of 64 and 40 days, respectively. Despite the rapid incubation periods, other strain-specified properties of many prion isolates--including the size of proteinase K-resistant PrPSc, the pattern of cerebral PrPSc deposition, and the conformational stability--were remarkably conserved upon serial passage in Tg(M109) mice. Our results demonstrate that expression of BVPrP is sufficient to engender enhanced susceptibility to a diverse range of prion isolates, suggesting that BVPrP may be a universal acceptor for prions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

Similar articles
-
Guinea Pig Prion Protein Supports Rapid Propagation of Bovine Spongiform Encephalopathy and Variant Creutzfeldt-Jakob Disease Prions.J Virol. 2016 Oct 14;90(21):9558-9569. doi: 10.1128/JVI.01106-16. Print 2016 Nov 1. J Virol. 2016. PMID: 27440899 Free PMC article.
-
Transmission barriers for bovine, ovine, and human prions in transgenic mice.J Virol. 2005 May;79(9):5259-71. doi: 10.1128/JVI.79.9.5259-5271.2005. J Virol. 2005. PMID: 15827140 Free PMC article.
-
Towards authentic transgenic mouse models of heritable PrP prion diseases.Acta Neuropathol. 2016 Oct;132(4):593-610. doi: 10.1007/s00401-016-1585-6. Epub 2016 Jun 28. Acta Neuropathol. 2016. PMID: 27350609 Free PMC article.
-
Insights into Mechanisms of Transmission and Pathogenesis from Transgenic Mouse Models of Prion Diseases.Methods Mol Biol. 2017;1658:219-252. doi: 10.1007/978-1-4939-7244-9_16. Methods Mol Biol. 2017. PMID: 28861793 Free PMC article. Review.
-
Transgenic models of prion disease.Arch Virol Suppl. 2000;(16):113-24. doi: 10.1007/978-3-7091-6308-5_10. Arch Virol Suppl. 2000. PMID: 11214913 Review.
Cited by
-
Chronic Wasting Disease (CWD) in Cervids and the Consequences of a Mutable Protein Conformation.ACS Omega. 2022 Apr 4;7(15):12474-12492. doi: 10.1021/acsomega.2c00155. eCollection 2022 Apr 19. ACS Omega. 2022. PMID: 35465121 Free PMC article. Review.
-
Gene-edited murine cell lines for propagation of chronic wasting disease prions.Sci Rep. 2019 Aug 1;9(1):11151. doi: 10.1038/s41598-019-47629-z. Sci Rep. 2019. PMID: 31371793 Free PMC article.
-
Genetically engineered cellular models of prion propagation.Cell Tissue Res. 2023 Apr;392(1):63-80. doi: 10.1007/s00441-022-03630-z. Epub 2022 May 18. Cell Tissue Res. 2023. PMID: 35581386 Review.
-
Full restoration of specific infectivity and strain properties from pure mammalian prion protein.PLoS Pathog. 2019 Mar 25;15(3):e1007662. doi: 10.1371/journal.ppat.1007662. eCollection 2019 Mar. PLoS Pathog. 2019. PMID: 30908557 Free PMC article.
-
G-quadruplexes within prion mRNA: the missing link in prion disease?Nucleic Acids Res. 2014 Aug;42(14):9327-33. doi: 10.1093/nar/gku559. Epub 2014 Jul 16. Nucleic Acids Res. 2014. PMID: 25030900 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- AG002132/AG/NIA NIH HHS/United States
- P01 AG002132/AG/NIA NIH HHS/United States
- R37 AG031220/AG/NIA NIH HHS/United States
- P01 AG010770/AG/NIA NIH HHS/United States
- CAPMC/ CIHR/Canada
- NS064173/NS/NINDS NIH HHS/United States
- P01 AG021601/AG/NIA NIH HHS/United States
- AG021601/AG/NIA NIH HHS/United States
- AG031220/AG/NIA NIH HHS/United States
- R00 NS064173/NS/NINDS NIH HHS/United States
- K99 AG042453/AG/NIA NIH HHS/United States
- AG010770/AG/NIA NIH HHS/United States
- K99 NS064173/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
