

Succinate is an inflammatory signal that induces IL-1β through HIF-1α
- PMID: 23535595
- PMCID: PMC4031686
- DOI: 10.1038/nature11986
Succinate is an inflammatory signal that induces IL-1β through HIF-1α
Abstract
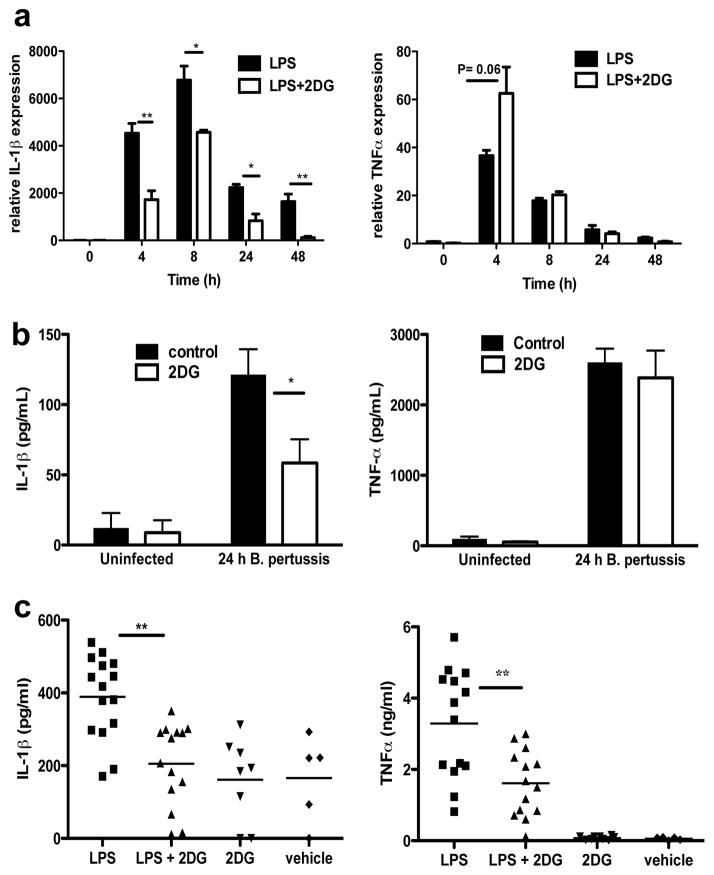
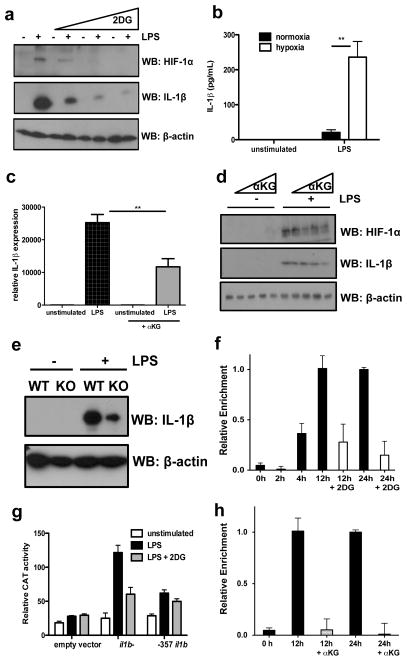
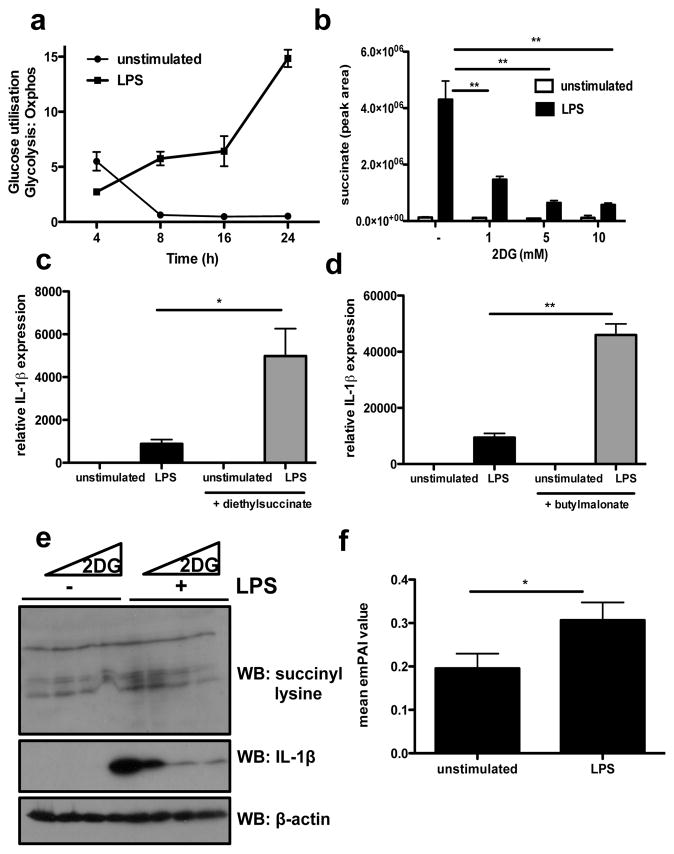
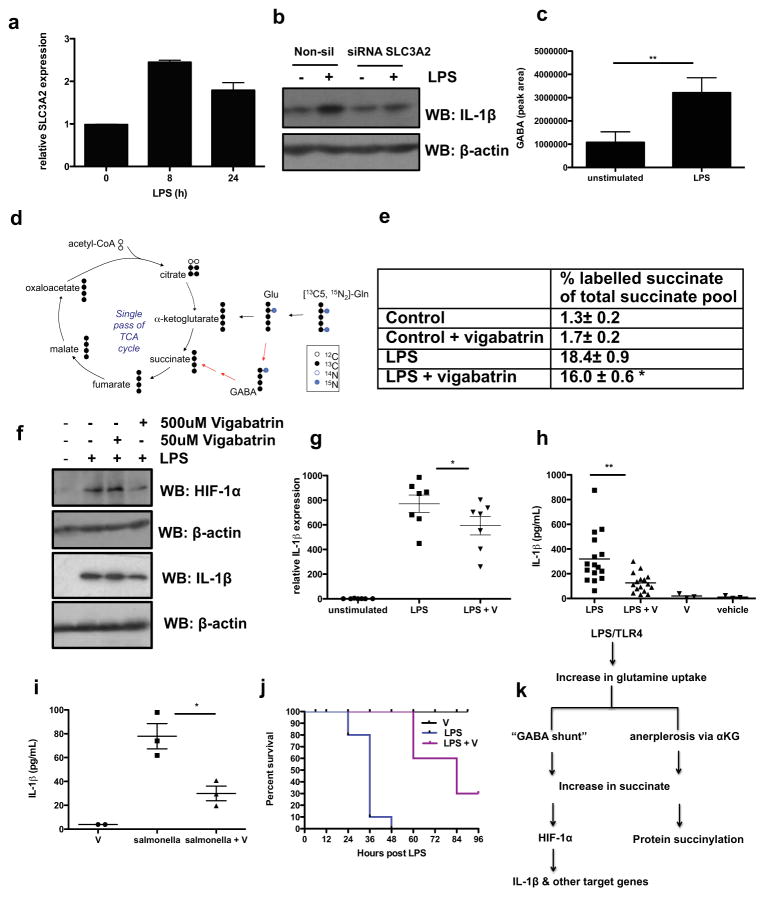
Macrophages activated by the Gram-negative bacterial product lipopolysaccharide switch their core metabolism from oxidative phosphorylation to glycolysis. Here we show that inhibition of glycolysis with 2-deoxyglucose suppresses lipopolysaccharide-induced interleukin-1β but not tumour-necrosis factor-α in mouse macrophages. A comprehensive metabolic map of lipopolysaccharide-activated macrophages shows upregulation of glycolytic and downregulation of mitochondrial genes, which correlates directly with the expression profiles of altered metabolites. Lipopolysaccharide strongly increases the levels of the tricarboxylic-acid cycle intermediate succinate. Glutamine-dependent anerplerosis is the principal source of succinate, although the 'GABA (γ-aminobutyric acid) shunt' pathway also has a role. Lipopolysaccharide-induced succinate stabilizes hypoxia-inducible factor-1α, an effect that is inhibited by 2-deoxyglucose, with interleukin-1β as an important target. Lipopolysaccharide also increases succinylation of several proteins. We therefore identify succinate as a metabolite in innate immune signalling, which enhances interleukin-1β production during inflammation.
Figures
Comment in
-
Agitation by suffocation: how hypoxia activates innate immunity via the Warburg effect.Cell Metab. 2013 Jun 4;17(6):814-815. doi: 10.1016/j.cmet.2013.05.016. Cell Metab. 2013. PMID: 23747241
-
mSphere of Influence: Host Metabolism Is an Integral Part of the Immune Response to Infectious Diseases.mSphere. 2021 Aug 25;6(4):e0024721. doi: 10.1128/mSphere.00247-21. Epub 2021 Jul 7. mSphere. 2021. PMID: 34232079 Free PMC article.
Similar articles
-
Agitation by suffocation: how hypoxia activates innate immunity via the Warburg effect.Cell Metab. 2013 Jun 4;17(6):814-815. doi: 10.1016/j.cmet.2013.05.016. Cell Metab. 2013. PMID: 23747241
-
Enhanced glycolysis and HIF-1α activation in adipose tissue macrophages sustains local and systemic interleukin-1β production in obesity.Sci Rep. 2020 Mar 27;10(1):5555. doi: 10.1038/s41598-020-62272-9. Sci Rep. 2020. PMID: 32221369 Free PMC article.
-
Succinate: a metabolic signal in inflammation.Trends Cell Biol. 2014 May;24(5):313-20. doi: 10.1016/j.tcb.2013.11.008. Epub 2013 Dec 19. Trends Cell Biol. 2014. PMID: 24361092 Review.
-
The miR-26a/SIRT6/HIF-1α axis regulates glycolysis and inflammatory responses in host macrophages during Mycobacterium tuberculosis infection.FEBS Lett. 2024 Oct;598(20):2592-2614. doi: 10.1002/1873-3468.15001. Epub 2024 Aug 18. FEBS Lett. 2024. PMID: 39155147
-
The Warburg effect then and now: from cancer to inflammatory diseases.Bioessays. 2013 Nov;35(11):965-73. doi: 10.1002/bies.201300084. Epub 2013 Sep 20. Bioessays. 2013. PMID: 24115022 Review.
Cited by
-
Branched-chain ketoacids derived from cancer cells modulate macrophage polarization and metabolic reprogramming.Front Immunol. 2022 Oct 13;13:966158. doi: 10.3389/fimmu.2022.966158. eCollection 2022. Front Immunol. 2022. PMID: 36311795 Free PMC article.
-
Reduction of host cell mitochondrial activity as Mycobacterium leprae's strategy to evade host innate immunity.Immunol Rev. 2021 May;301(1):193-208. doi: 10.1111/imr.12962. Epub 2021 Apr 28. Immunol Rev. 2021. PMID: 33913182 Free PMC article. Review.
-
Food Fight: Role of Itaconate and Other Metabolites in Antimicrobial Defense.Cell Metab. 2016 Sep 13;24(3):379-387. doi: 10.1016/j.cmet.2016.08.013. Cell Metab. 2016. PMID: 27626199 Free PMC article. Review.
-
Staphylococcus aureus induces an itaconate-dominated immunometabolic response that drives biofilm formation.Nat Commun. 2021 Mar 3;12(1):1399. doi: 10.1038/s41467-021-21718-y. Nat Commun. 2021. PMID: 33658521 Free PMC article.
-
Cadmium-mediated lung injury is exacerbated by the persistence of classically activated macrophages.J Biol Chem. 2020 Nov 13;295(46):15754-15766. doi: 10.1074/jbc.RA120.013632. Epub 2020 Sep 11. J Biol Chem. 2020. PMID: 32917723 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
