

RNF4-dependent hybrid SUMO-ubiquitin chains are signals for RAP80 and thereby mediate the recruitment of BRCA1 to sites of DNA damage
- PMID: 23211528
- PMCID: PMC4131685
- DOI: 10.1126/scisignal.2003485
RNF4-dependent hybrid SUMO-ubiquitin chains are signals for RAP80 and thereby mediate the recruitment of BRCA1 to sites of DNA damage
Abstract
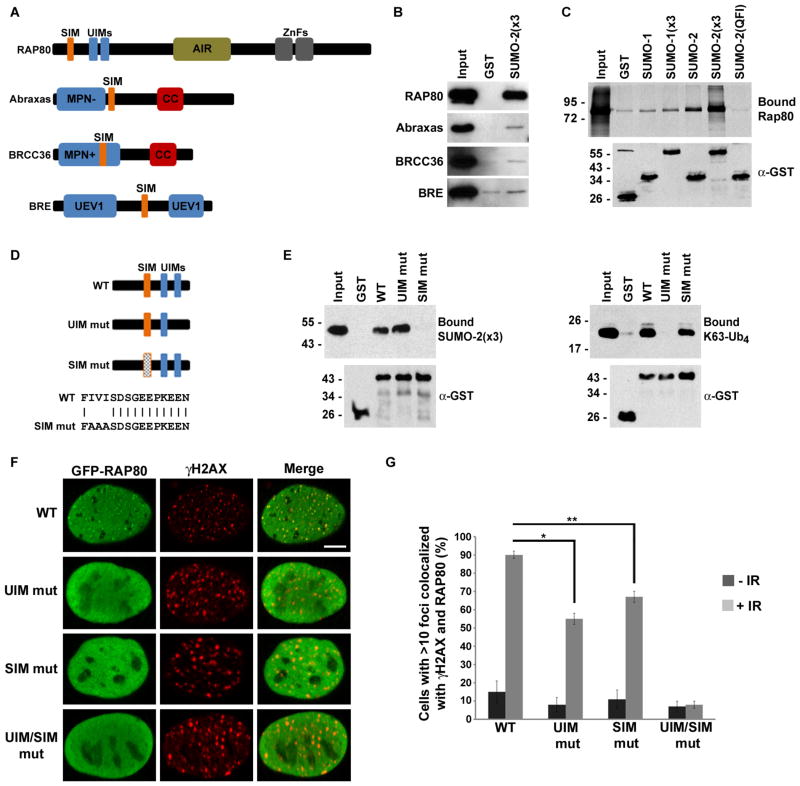
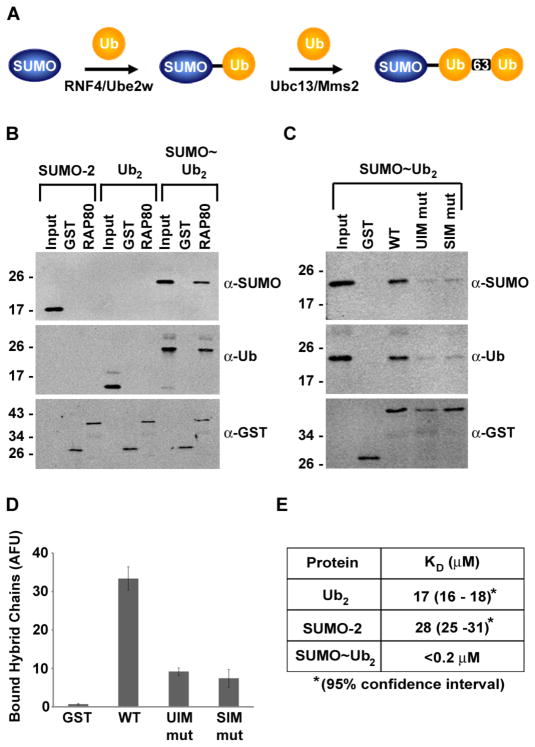
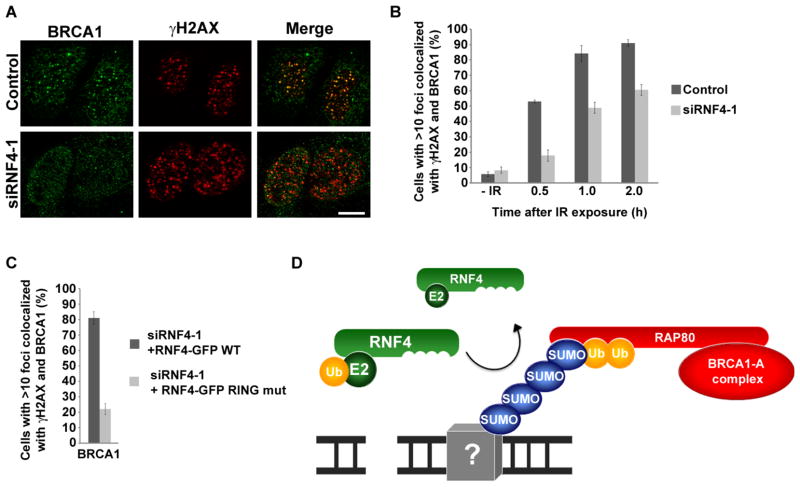
The DNA repair function of the breast cancer susceptibility protein BRCA1 depends in part on its interaction with RAP80, which targets BRCA1 to DNA double-strand breaks (DSBs) through recognition of K63-linked polyubiquitin chains. The localization of BRCA1 to DSBs also requires sumoylation. We demonstrated that, in addition to having ubiquitin-interacting motifs, RAP80 also contains a SUMO-interacting motif (SIM) that is critical for recruitment to DSBs. In combination with the ubiquitin-binding activity of RAP80, this SIM enabled RAP80 to bind with nanomolar affinity to hybrid chains consisting of ubiquitin conjugated to SUMO. Furthermore, RNF4, a SUMO-targeted ubiquitin E3 ligase that synthesizes hybrid SUMO-ubiquitin chains, localized to DSBs and was critical for the recruitment of RAP80 and BRCA1 to sites of DNA damage. Our findings, therefore, connect ubiquitin- and SUMO-dependent DSB recognition, revealing that RNF4-synthesized hybrid SUMO-ubiquitin chains are recognized by RAP80 to promote BRCA1 recruitment and DNA repair.
Conflict of interest statement
Figures

Similar articles
-
Rap80 protein recruitment to DNA double-strand breaks requires binding to both small ubiquitin-like modifier (SUMO) and ubiquitin conjugates.J Biol Chem. 2012 Jul 20;287(30):25510-9. doi: 10.1074/jbc.M112.374116. Epub 2012 Jun 11. J Biol Chem. 2012. PMID: 22689573 Free PMC article.
-
Ataxin-3 consolidates the MDC1-dependent DNA double-strand break response by counteracting the SUMO-targeted ubiquitin ligase RNF4.EMBO J. 2017 Apr 13;36(8):1066-1083. doi: 10.15252/embj.201695151. Epub 2017 Mar 8. EMBO J. 2017. PMID: 28275011 Free PMC article.
-
Molecular Basis for Phosphorylation-dependent SUMO Recognition by the DNA Repair Protein RAP80.J Biol Chem. 2016 Feb 26;291(9):4417-28. doi: 10.1074/jbc.M115.705061. Epub 2015 Dec 30. J Biol Chem. 2016. PMID: 26719330 Free PMC article.
-
RAP80, ubiquitin and SUMO in the DNA damage response.J Mol Med (Berl). 2017 Aug;95(8):799-807. doi: 10.1007/s00109-017-1561-1. Epub 2017 Jul 5. J Mol Med (Berl). 2017. PMID: 28681078 Free PMC article. Review.
-
RAP80 and RNF8, key players in the recruitment of repair proteins to DNA damage sites.Cancer Lett. 2008 Nov 28;271(2):179-90. doi: 10.1016/j.canlet.2008.04.046. Epub 2008 Jun 11. Cancer Lett. 2008. PMID: 18550271 Free PMC article. Review.
Cited by
-
Crosstalk between SUMOylation and other post-translational modifications in breast cancer.Cell Mol Biol Lett. 2024 Aug 10;29(1):107. doi: 10.1186/s11658-024-00624-3. Cell Mol Biol Lett. 2024. PMID: 39127633 Free PMC article. Review.
-
SUMO and the DNA damage response.Biochem Soc Trans. 2024 Apr 24;52(2):773-792. doi: 10.1042/BST20230862. Biochem Soc Trans. 2024. PMID: 38629643 Free PMC article. Review.
-
RNF4 sustains Myc-driven tumorigenesis by facilitating DNA replication.J Clin Invest. 2024 Mar 26;134(10):e167419. doi: 10.1172/JCI167419. J Clin Invest. 2024. PMID: 38530355 Free PMC article.
-
SUMOylation controls Hu antigen R posttranscriptional activity in liver cancer.Cell Rep. 2024 Mar 26;43(3):113924. doi: 10.1016/j.celrep.2024.113924. Epub 2024 Mar 18. Cell Rep. 2024. PMID: 38507413 Free PMC article.
-
Repression of the SUMO-conjugating enzyme UBC9 is associated with lowered double minutes and reduced tumor progression.Cancer Biol Ther. 2024 Dec 31;25(1):2323768. doi: 10.1080/15384047.2024.2323768. Epub 2024 Mar 11. Cancer Biol Ther. 2024. PMID: 38465861 Free PMC article.
References
-
- Bensimon A, Schmidt A, Ziv Y, Elkon R, Wang SY, Chen DJ, Aebersold R, Shiloh Y. ATM-dependent and -independent dynamics of the nuclear phosphoproteome after DNA damage. Sci Signal. 2010;3:rs3. - PubMed
-
- Rupnik A, Lowndes NF, Grenon M. MRN and the race to the break. Chromosoma. 2010;119:115–135. - PubMed
-
- Bekker-Jensen S, Mailand N. The ubiquitin- and SUMO-dependent signaling response to DNA double-strand breaks. FEBS Lett. 2011;585:2914–2919. - PubMed
-
- Bergink S, Jentsch S. Principles of ubiquitin and SUMO modifications in DNA repair. Nature. 2009;458:461–467. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
