

Tobacco calmodulin-like protein provides secondary defense by binding to and directing degradation of virus RNA silencing suppressors
- PMID: 22665793
- PMCID: PMC3382489
- DOI: 10.1073/pnas.1201628109
Tobacco calmodulin-like protein provides secondary defense by binding to and directing degradation of virus RNA silencing suppressors
Abstract
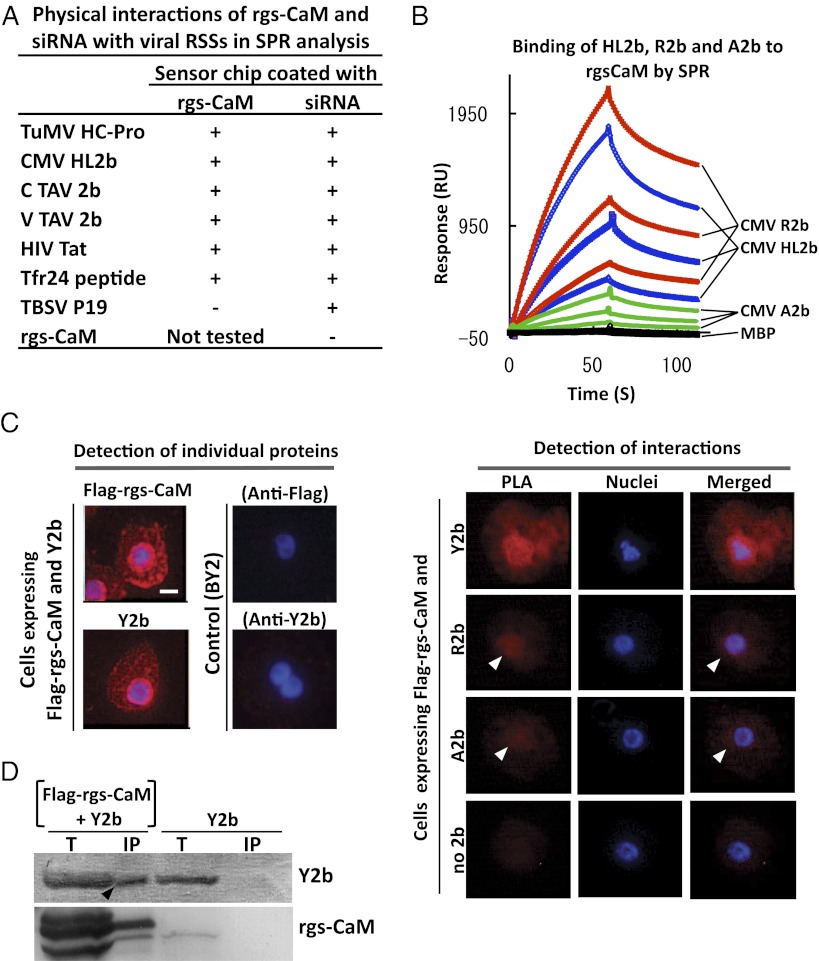
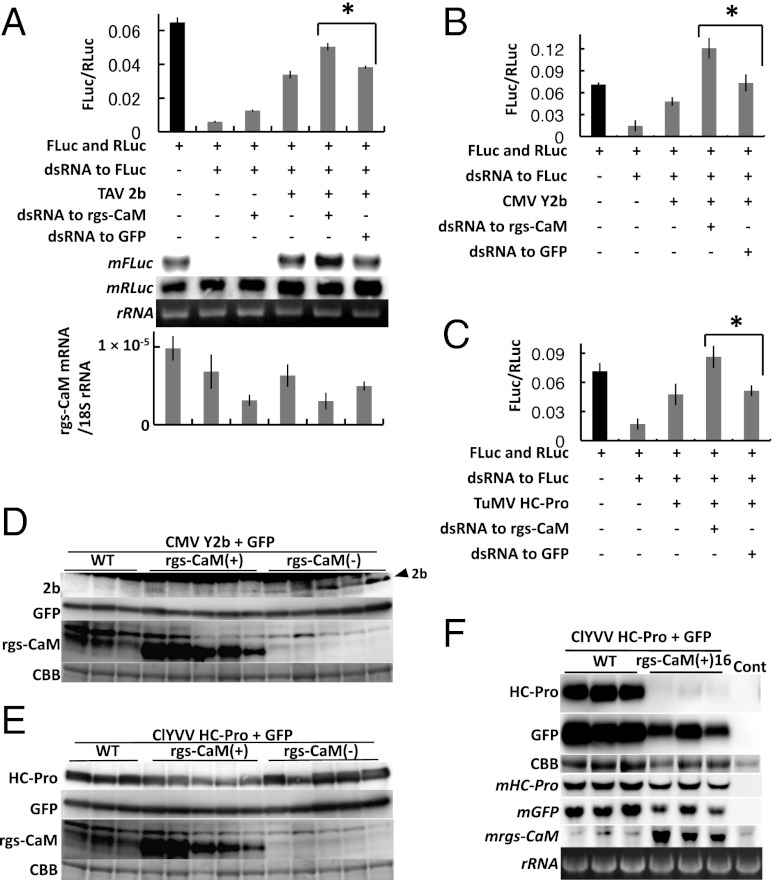
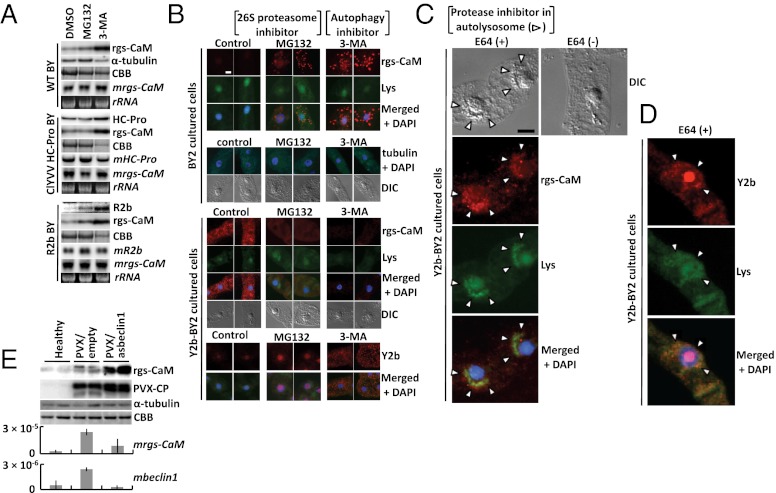
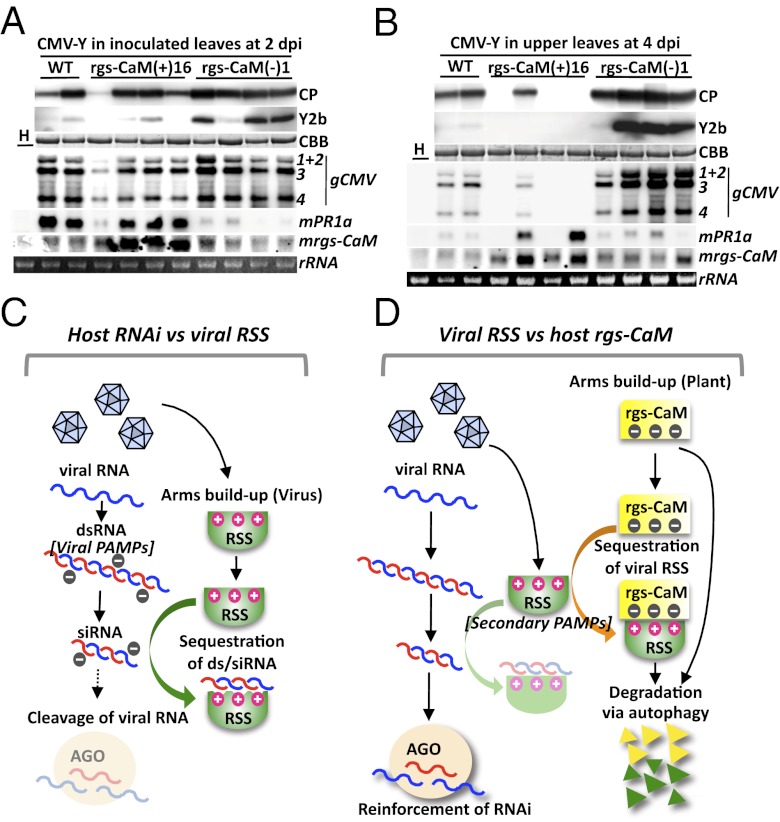
RNA silencing (RNAi) induced by virus-derived double-stranded RNA (dsRNA), which is in a sense regarded as a pathogen-associated molecular pattern (PAMP) of viruses, is a general plant defense mechanism. To counteract this defense, plant viruses express RNA silencing suppressors (RSSs), many of which bind to dsRNA and attenuate RNAi. We showed that the tobacco calmodulin-like protein, rgs-CaM, counterattacked viral RSSs by binding to their dsRNA-binding domains and sequestering them from inhibiting RNAi. Autophagy-like protein degradation seemed to operate to degrade RSSs with the sacrifice of rgs-CaM. These RSSs could thus be regarded as secondary viral PAMPs. This study uncovered a unique defense system in which an rgs-CaM-mediated countermeasure against viral RSSs enhanced host antiviral RNAi in tobacco.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Wound-induced rgs-CaM gets ready for counterresponse to an early stage of viral infection.Plant Signal Behav. 2012 Dec;7(12):1548-51. doi: 10.4161/psb.22369. Epub 2012 Oct 16. Plant Signal Behav. 2012. PMID: 23073002 Free PMC article.
-
rgs-CaM Detects and Counteracts Viral RNA Silencing Suppressors in Plant Immune Priming.J Virol. 2017 Sep 12;91(19):e00761-17. doi: 10.1128/JVI.00761-17. Print 2017 Oct 1. J Virol. 2017. PMID: 28724770 Free PMC article.
-
Double-stranded RNA binding may be a general plant RNA viral strategy to suppress RNA silencing.J Virol. 2006 Jun;80(12):5747-56. doi: 10.1128/JVI.01963-05. J Virol. 2006. PMID: 16731914 Free PMC article.
-
Viral RNA silencing suppressors (RSS): novel strategy of viruses to ablate the host RNA interference (RNAi) defense system.Virus Res. 2011 Jan;155(1):1-9. doi: 10.1016/j.virusres.2010.10.003. Epub 2010 Oct 14. Virus Res. 2011. PMID: 20951748 Free PMC article. Review.
-
The dual edge of RNA silencing suppressors in the virus-host interactions.Curr Opin Virol. 2016 Apr;17:39-44. doi: 10.1016/j.coviro.2015.12.002. Epub 2016 Jan 21. Curr Opin Virol. 2016. PMID: 26802204 Review.
Cited by
-
Two amino acids near the N-terminus of Cucumber mosaic virus 2b play critical roles in the suppression of RNA silencing and viral infectivity.Mol Plant Pathol. 2016 Feb;17(2):173-83. doi: 10.1111/mpp.12270. Epub 2015 Jun 7. Mol Plant Pathol. 2016. PMID: 25893424 Free PMC article.
-
Autophagy-virus interplay in plants: from antiviral recognition to proviral manipulation.Mol Plant Pathol. 2019 Sep;20(9):1211-1216. doi: 10.1111/mpp.12852. Epub 2019 Aug 9. Mol Plant Pathol. 2019. PMID: 31397085 Free PMC article. Review.
-
Molecular Regulation of Host Defense Responses Mediated by Biological Anti-TMV Agent Ningnanmycin.Viruses. 2019 Sep 3;11(9):815. doi: 10.3390/v11090815. Viruses. 2019. PMID: 31484426 Free PMC article.
-
Autophagy Inhibits Intercellular Transport of Citrus Leaf Blotch Virus by Targeting Viral Movement Protein.Viruses. 2021 Oct 30;13(11):2189. doi: 10.3390/v13112189. Viruses. 2021. PMID: 34834995 Free PMC article.
-
Sense transgene-induced post-transcriptional gene silencing in tobacco compromises the splicing of endogenous counterpart genes.PLoS One. 2014 Feb 21;9(2):e87869. doi: 10.1371/journal.pone.0087869. eCollection 2014. PLoS One. 2014. PMID: 24586294 Free PMC article.
References
-
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
-
- Chisholm ST, Coaker G, Day B, Staskawicz BJ. Host-microbe interactions: Shaping the evolution of the plant immune response. Cell. 2006;124:803–814. - PubMed
-
- Boller T, Felix G. A renaissance of elicitors: Perception of microbe-associated molecular patterns and danger signals by pattern-recognition receptors. Annu Rev Plant Biol. 2009;60:379–406. - PubMed
-
- Alfano JR, Collmer A. Type III secretion system effector proteins: Double agents in bacterial disease and plant defense. Annu Rev Phytopathol. 2004;42:385–414. - PubMed
-
- Abramovitch RB, Martin GB. Strategies used by bacterial pathogens to suppress plant defenses. Curr Opin Plant Biol. 2004;7:356–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
