

Phosphoproteomic analysis reveals that PP4 dephosphorylates KAP-1 impacting the DNA damage response
- PMID: 22491012
- PMCID: PMC3364739
- DOI: 10.1038/emboj.2012.86
Phosphoproteomic analysis reveals that PP4 dephosphorylates KAP-1 impacting the DNA damage response
Abstract
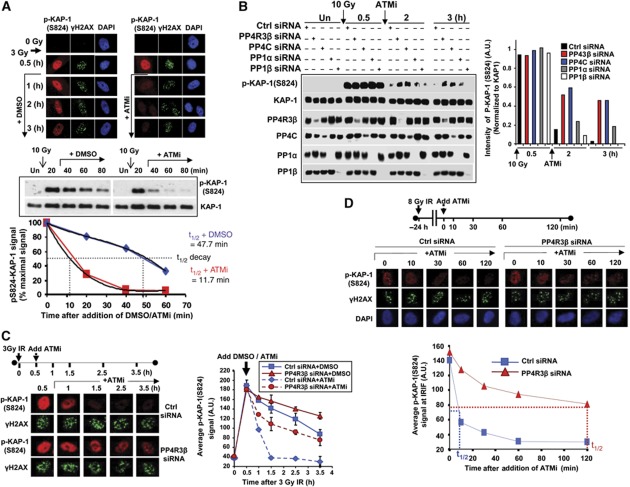
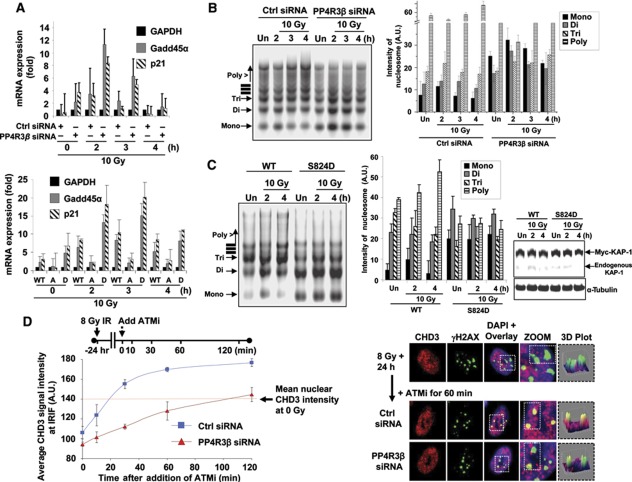
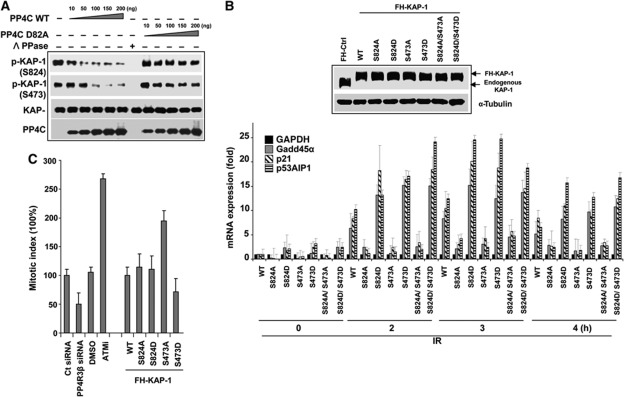
Protein phosphatase PP4C has been implicated in the DNA damage response (DDR), but its substrates in DDR remain largely unknown. We devised a novel proteomic strategy for systematic identification of proteins dephosphorylated by PP4C and identified KRAB-domain-associated protein 1 (KAP-1) as a substrate. Ionizing radiation leads to phosphorylation of KAP-1 at S824 (via ATM) and at S473 (via CHK2). A PP4C/R3β complex interacts with KAP-1 and silencing this complex leads to persistence of phospho-S824 and phospho-S473. We identify a new role for KAP-1 in DDR by showing that phosphorylation of S473 impacts the G2/M checkpoint. Depletion of PP4R3β or expression of the phosphomimetic KAP-1 S473 mutant (S473D) leads to a prolonged G2/M checkpoint. Phosphorylation of S824 is necessary for repair of heterochromatic DNA lesions and similar to cells expressing phosphomimetic KAP-1 S824 mutant (S824D), or PP4R3β-silenced cells, display prolonged relaxation of chromatin with release of chromatin remodelling protein CHD3. Our results define a new role for PP4-mediated dephosphorylation in the DDR, including the regulation of a previously undescribed function of KAP-1 in checkpoint response.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




Comment in
-
When breaking is bad but repair is worse.Mol Cell. 2014 May 8;54(3):332-3. doi: 10.1016/j.molcel.2014.04.021. Mol Cell. 2014. PMID: 24813711
Similar articles
-
Protein phosphatase PP4 is involved in NHEJ-mediated repair of DNA double-strand breaks.Cell Cycle. 2012 Jul 15;11(14):2643-9. doi: 10.4161/cc.20957. Epub 2012 Jul 15. Cell Cycle. 2012. PMID: 22732494
-
Kruppel-associated Box (KRAB)-associated co-repressor (KAP-1) Ser-473 phosphorylation regulates heterochromatin protein 1β (HP1-β) mobilization and DNA repair in heterochromatin.J Biol Chem. 2012 Aug 10;287(33):28122-31. doi: 10.1074/jbc.M112.368381. Epub 2012 Jun 18. J Biol Chem. 2012. PMID: 22715096 Free PMC article.
-
A PP4 phosphatase complex dephosphorylates RPA2 to facilitate DNA repair via homologous recombination.Nat Struct Mol Biol. 2010 Mar;17(3):365-72. doi: 10.1038/nsmb.1769. Epub 2010 Feb 14. Nat Struct Mol Biol. 2010. PMID: 20154705 Free PMC article.
-
Emerging role of protein phosphatases changes the landscape of phospho-signaling in DNA damage response.DNA Repair (Amst). 2015 Aug;32:58-65. doi: 10.1016/j.dnarep.2015.04.014. Epub 2015 May 1. DNA Repair (Amst). 2015. PMID: 25956864 Review.
-
The influence of heterochromatin on DNA double strand break repair: Getting the strong, silent type to relax.DNA Repair (Amst). 2010 Dec 10;9(12):1273-82. doi: 10.1016/j.dnarep.2010.09.013. Epub 2010 Oct 30. DNA Repair (Amst). 2010. PMID: 21036673 Review.
Cited by
-
A role for protein phosphatase 4 in regulating non-homologous end-joining.Cell Cycle. 2012 Oct 1;11(19):3534-5. doi: 10.4161/cc.22040. Epub 2012 Sep 5. Cell Cycle. 2012. PMID: 22951540 Free PMC article. No abstract available.
-
Double-strand break repair-adox: Restoration of suppressed double-strand break repair during mitosis induces genomic instability.Cancer Sci. 2014 Dec;105(12):1519-25. doi: 10.1111/cas.12551. Epub 2014 Nov 5. Cancer Sci. 2014. PMID: 25287622 Free PMC article. Review.
-
ATM/Wip1 activities at chromatin control Plk1 re-activation to determine G2 checkpoint duration.EMBO J. 2017 Jul 14;36(14):2161-2176. doi: 10.15252/embj.201696082. Epub 2017 Jun 12. EMBO J. 2017. PMID: 28607002 Free PMC article.
-
Dephosphorylation of DBC1 by Protein Phosphatase 4 Is Important for p53-Mediated Cellular Functions.Mol Cells. 2015 Aug;38(8):697-704. doi: 10.14348/molcells.2015.0066. Epub 2015 Jul 21. Mol Cells. 2015. PMID: 26194823 Free PMC article.
-
WIP1 Promotes Homologous Recombination and Modulates Sensitivity to PARP Inhibitors.Cells. 2019 Oct 15;8(10):1258. doi: 10.3390/cells8101258. Cells. 2019. PMID: 31619012 Free PMC article.
References
-
- Ahn JY, Schwarz JK, Piwnica-Worms H, Canman CE (2000) Threonine 68 phosphorylation by ataxia telangiectasia mutated is required for efficient activation of Chk2 in response to ionizing radiation. Cancer Res 60: 5934–5936 - PubMed
-
- Arroyo JD, Lee GM, Hahn WC (2008) Liprin alpha1 interacts with PP2A B56gamma. Cell Cycle 7: 525–532 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
