

EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency
- PMID: 22473208
- PMCID: PMC3343464
- DOI: 10.1038/emboj.2012.63
EBV and human microRNAs co-target oncogenic and apoptotic viral and human genes during latency
Abstract
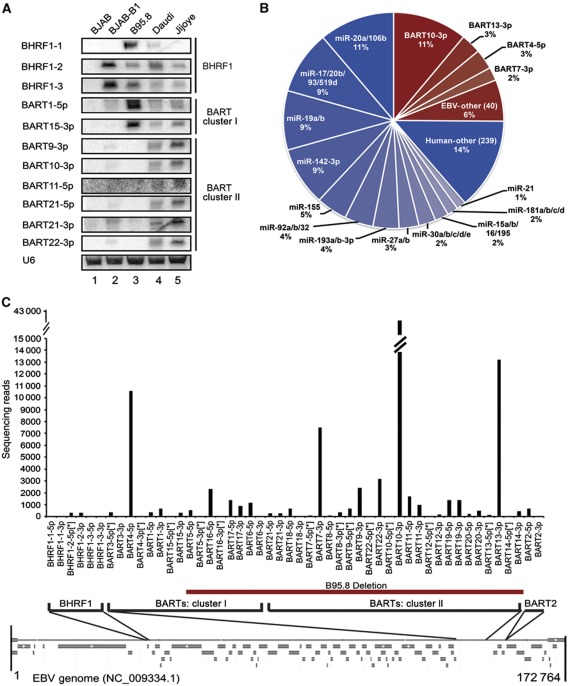
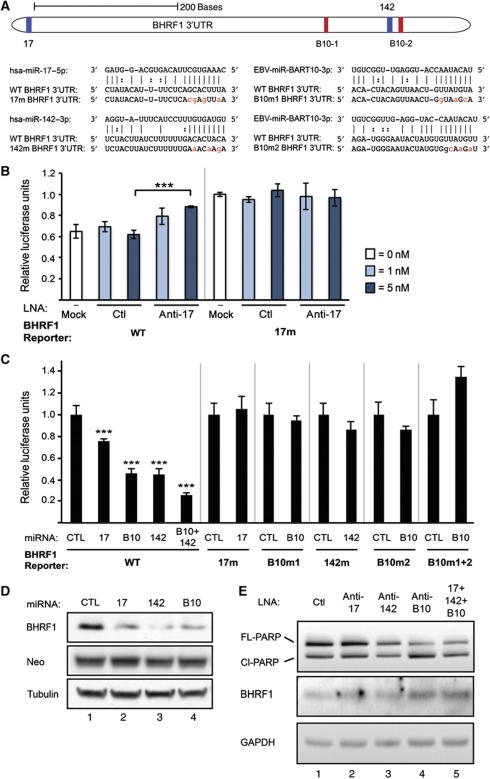
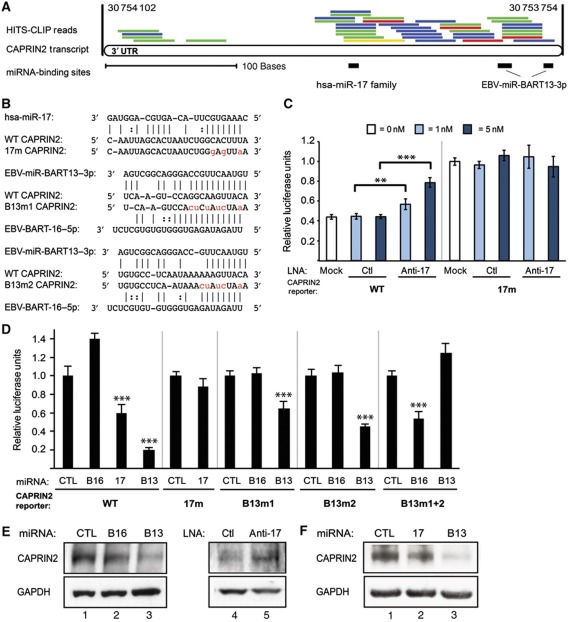
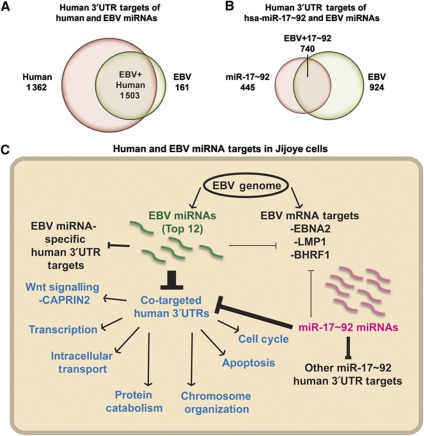
Epstein-Barr virus (EBV) controls gene expression to transform human B cells and maintain viral latency. High-throughput sequencing and crosslinking immunoprecipitation (HITS-CLIP) identified mRNA targets of 44 EBV and 310 human microRNAs (miRNAs) in Jijoye (Latency III) EBV-transformed B cells. While 25% of total cellular miRNAs are viral, only three viral mRNAs, all latent transcripts, are targeted. Thus, miRNAs do not control the latent/lytic switch by targeting EBV lytic genes. Unexpectedly, 90% of the 1664 human 3'-untranslated regions targeted by the 12 most abundant EBV miRNAs are also targeted by human miRNAs via distinct binding sites. Half of these are targets of the oncogenic miR-17∼92 miRNA cluster and associated families, including mRNAs that regulate transcription, apoptosis, Wnt signalling, and the cell cycle. Reporter assays confirmed the functionality of several EBV and miR-17 family miRNA-binding sites in EBV latent membrane protein 1 (LMP1), EBV BHRF1, and host CAPRIN2 mRNAs. Our extensive list of EBV and human miRNA targets implicates miRNAs in the control of EBV latency and illuminates viral miRNA function in general.
Conflict of interest statement
The authors delcare that they have no conflict of interest.
Figures


Similar articles
-
MicroRNA miR-BART20-5p stabilizes Epstein-Barr virus latency by directly targeting BZLF1 and BRLF1.J Virol. 2014 Aug;88(16):9027-37. doi: 10.1128/JVI.00721-14. Epub 2014 Jun 4. J Virol. 2014. PMID: 24899173 Free PMC article.
-
A viral microRNA cluster strongly potentiates the transforming properties of a human herpesvirus.PLoS Pathog. 2011 Feb;7(2):e1001294. doi: 10.1371/journal.ppat.1001294. Epub 2011 Feb 17. PLoS Pathog. 2011. PMID: 21379335 Free PMC article.
-
Epstein-Barr Virus miR-BHRF1-3 Targets the BZLF1 3'UTR and Regulates the Lytic Cycle.J Virol. 2022 Feb 23;96(4):e0149521. doi: 10.1128/JVI.01495-21. Epub 2021 Dec 8. J Virol. 2022. PMID: 34878852 Free PMC article.
-
Role of Viral and Host microRNAs in Immune Regulation of Epstein-Barr Virus-Associated Diseases.Front Immunol. 2020 Mar 3;11:367. doi: 10.3389/fimmu.2020.00367. eCollection 2020. Front Immunol. 2020. PMID: 32194570 Free PMC article. Review.
-
The role of microRNAs in Epstein-Barr virus latency and lytic reactivation.Microbes Infect. 2011 Dec;13(14-15):1156-67. doi: 10.1016/j.micinf.2011.07.007. Epub 2011 Jul 28. Microbes Infect. 2011. PMID: 21835261 Free PMC article. Review.
Cited by
-
MicroRNAs as mediators of viral evasion of the immune system.Nat Immunol. 2013 Mar;14(3):205-10. doi: 10.1038/ni.2537. Epub 2013 Feb 15. Nat Immunol. 2013. PMID: 23416678 Free PMC article. Review.
-
Virus-encoded microRNAs: an overview and a look to the future.PLoS Pathog. 2012 Dec;8(12):e1003018. doi: 10.1371/journal.ppat.1003018. Epub 2012 Dec 20. PLoS Pathog. 2012. PMID: 23308061 Free PMC article. Review.
-
The Function and Therapeutic Potential of Epstein-Barr Virus-Encoded MicroRNAs in Cancer.Mol Ther Nucleic Acids. 2019 Sep 6;17:657-668. doi: 10.1016/j.omtn.2019.07.002. Epub 2019 Jul 15. Mol Ther Nucleic Acids. 2019. PMID: 31400608 Free PMC article. Review.
-
Nascent Transcriptomics Reveal Cellular Prolytic Factors Upregulated Upstream of the Latent-to-Lytic Switch Protein of Epstein-Barr Virus.J Virol. 2020 Mar 17;94(7):e01966-19. doi: 10.1128/JVI.01966-19. Print 2020 Mar 17. J Virol. 2020. PMID: 31941784 Free PMC article.
-
Reciprocal inhibition between intracellular antiviral signaling and the RNAi machinery in mammalian cells.Cell Host Microbe. 2013 Oct 16;14(4):435-45. doi: 10.1016/j.chom.2013.09.002. Epub 2013 Sep 26. Cell Host Microbe. 2013. PMID: 24075860 Free PMC article.
References
-
- Aerbajinai W, Lee YT, Wojda U, Barr VA, Miller JL (2004) Cloning and characterization of a gene expressed during terminal differentiation that encodes a novel inhibitor of growth. J Biol Chem 279: 1916–1921 - PubMed
-
- Al-Mozaini M, Bodelon G, Karstegl CE, Jin B, Al-Ahdal M, Farrell PJ (2009) Epstein-Barr virus BART gene expression. J Gen Virol 90: 307–316 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
