

Evidence of Fe3+ interaction with the plug domain of the outer membrane transferrin receptor protein of Neisseria gonorrhoeae: implications for Fe transport
- PMID: 22399131
- PMCID: PMC3391718
- DOI: 10.1039/c2mt20037f
Evidence of Fe3+ interaction with the plug domain of the outer membrane transferrin receptor protein of Neisseria gonorrhoeae: implications for Fe transport
Abstract
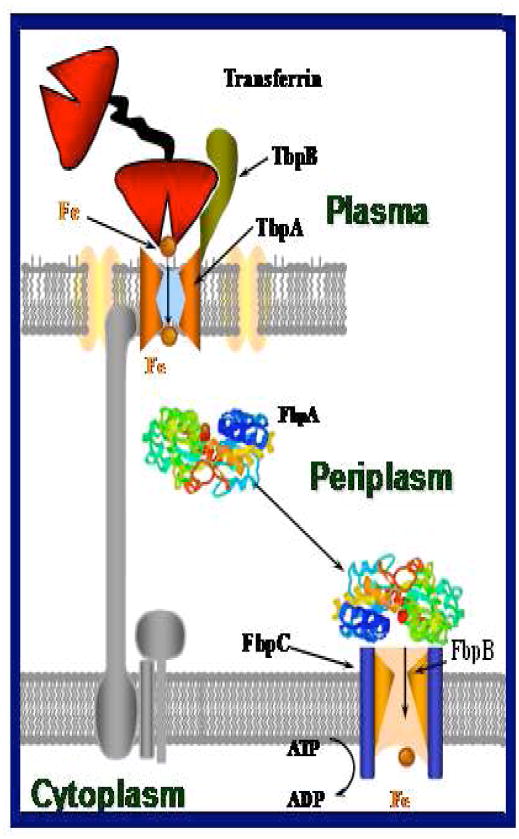
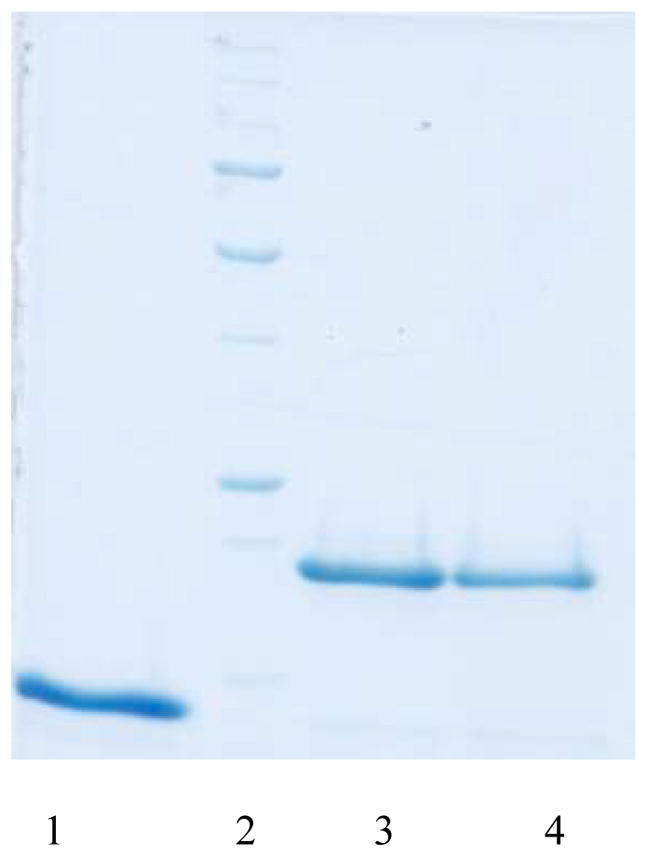
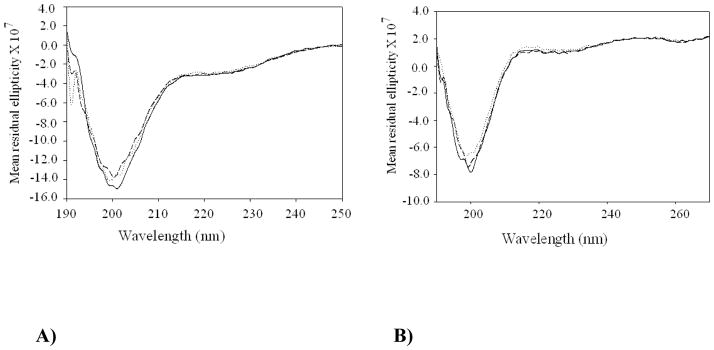
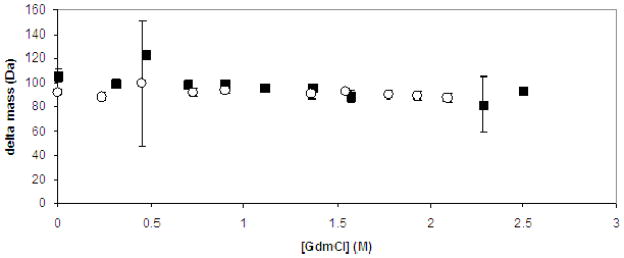
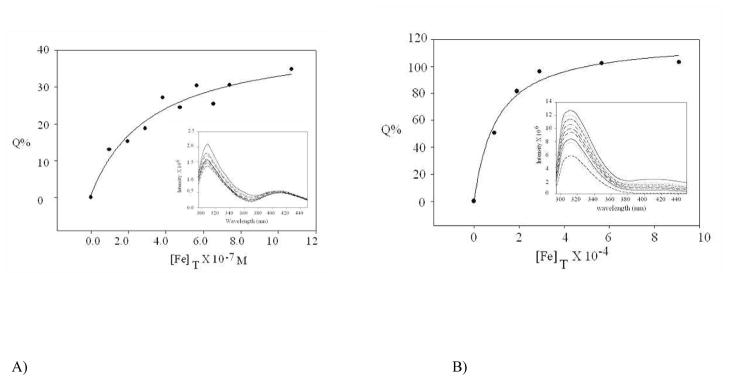
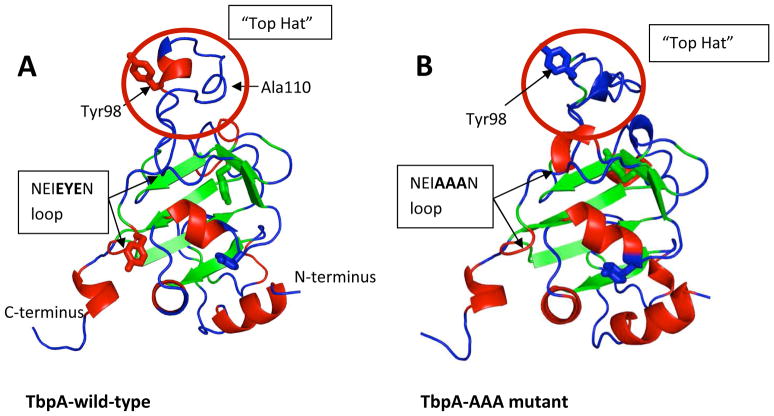
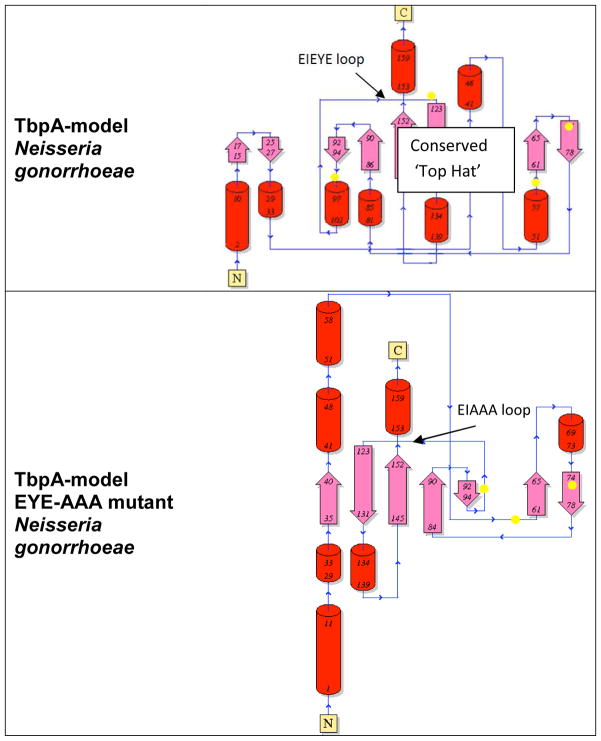
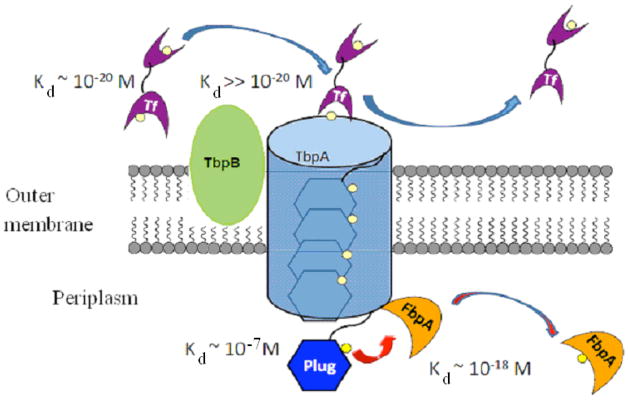
Neisseria gonorrhoeae is an obligate pathogen that hijacks iron from the human iron transport protein, holo-transferrin (Fe(2)-Tf), by expressing TonB-dependent outer membrane receptor proteins, TbpA and TbpB. Homologous to other TonB-dependent outer membrane transporters, TbpA is thought to consist of a β-barrel with an N-terminal plug domain. Previous reports by our laboratories show that the sequence EIEYE in the plug domain is highly conserved among various bacterial species that express TbpA and plays a crucial role in iron utilization for gonococci. We hypothesize that this highly conserved EIEYE sequence in the TbpA plug, rich in hard oxygen donor groups, binds with Fe(3+) through the transport process across the outer membrane through the β-barrel. Sequestration of Fe(3+) by the TbpA-plug supports the paradigm that the ferric iron must always remain chelated and controlled throughout the transport process. In order to test this hypothesis here we describe the ability of both the recombinant wild-type plug, and three small peptides that encompass the sequence EIEYE of the plug, to bind Fe(3+). This is the first report of the expression/isolation of the recombinant wild-type TbpA plug. Although CD and SUPREX spectroscopies suggest that a non-native structure is observed for the recombinant plug, fluorescence quenching titrations indicate that the wild-type recombinant TbpA plug binds Fe (3+) with a conditional log K(d) = 7 at pH 7.5, with no evidence of binding at pH 6.3. A recombinant TbpA plug with mutated sequence (NEIEYEN → NEIAAAN) shows no evidence of Fe(3+) binding under our experimental set up. Interestingly, in silico modeling with the wild-type plug also predicts a flexible loop structure for the EIEYE sequence under native conditions which once again supports the Fe(3+) binding hypothesis. These in vitro observations are consistent with the hypothesis that the EIEYE sequence in the wild-type TbpA plug binds Fe(3+) during the outer membrane transport process in vivo.
This journal is © The Royal Society of Chemistry 2012
Figures


Similar articles
-
Identification of TbpA residues required for transferrin-iron utilization by Neisseria gonorrhoeae.Infect Immun. 2008 May;76(5):1960-9. doi: 10.1128/IAI.00020-08. Epub 2008 Mar 17. Infect Immun. 2008. PMID: 18347046 Free PMC article.
-
Hijacking transferrin bound iron: protein-receptor interactions involved in iron transport in N. gonorrhoeae.Metallomics. 2009;1(3):249-55. doi: 10.1039/b902860a. Metallomics. 2009. PMID: 20161024 Free PMC article.
-
Determination of surface-exposed, functional domains of gonococcal transferrin-binding protein A.Infect Immun. 2004 Mar;72(3):1775-85. doi: 10.1128/IAI.72.3.1775-1785.2004. Infect Immun. 2004. PMID: 14977987 Free PMC article.
-
Molecular mechanism of ferricsiderophore passage through the outer membrane receptor proteins of Escherichia coli.Biometals. 2007 Jun;20(3-4):263-74. doi: 10.1007/s10534-006-9060-9. Epub 2006 Dec 22. Biometals. 2007. PMID: 17186377 Review.
-
Mechanisms of iron acquisition by the human pathogens Neisseria meningitidis and Neisseria gonorrhoeae.Front Biosci. 2003 Sep 1;8:d1186-218. doi: 10.2741/1133. Front Biosci. 2003. PMID: 12957813 Review.
Cited by
-
Bordetella pertussis FbpA binds both unchelated iron and iron siderophore complexes.Biochemistry. 2014 Jun 24;53(24):3952-60. doi: 10.1021/bi5002823. Epub 2014 Jun 13. Biochemistry. 2014. PMID: 24873326 Free PMC article.
-
Ferroptosis: A New Road towards Cancer Management.Molecules. 2022 Mar 25;27(7):2129. doi: 10.3390/molecules27072129. Molecules. 2022. PMID: 35408533 Free PMC article. Review.
-
Comparative Genomics of the Genus Porphyromonas Identifies Adaptations for Heme Synthesis within the Prevalent Canine Oral Species Porphyromonas cangingivalis.Genome Biol Evol. 2015 Nov 13;7(12):3397-413. doi: 10.1093/gbe/evv220. Genome Biol Evol. 2015. PMID: 26568374 Free PMC article.
-
The transferrin-iron import system from pathogenic Neisseria species.Mol Microbiol. 2012 Oct;86(2):246-57. doi: 10.1111/mmi.12002. Epub 2012 Sep 7. Mol Microbiol. 2012. PMID: 22957710 Free PMC article. Review.
-
Structural Basis for Evasion of Nutritional Immunity by the Pathogenic Neisseriae.Front Microbiol. 2020 Jan 10;10:2981. doi: 10.3389/fmicb.2019.02981. eCollection 2019. Front Microbiol. 2020. PMID: 31998268 Free PMC article. Review.
References
-
- Briat JF. J Gen Microbiol. 1992;138:2475–2483. - PubMed
-
- Griffiths E. Iron and Infection: molecular physiology and clinical aspects. :69–137.
-
- Imbert M, Blondeau R. Curr Microbiol. 1998;37:64–66. - PubMed
-
- Ratledge C, Dover LG. Annu Rev Microbiol. 2000;54:881–941. - PubMed
-
- Weinberg ED. Perspect Biol Med. 1997;40:578–583. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
