

Lens epithelium-derived growth factor fusion proteins redirect HIV-1 DNA integration
- PMID: 20133638
- PMCID: PMC2840313
- DOI: 10.1073/pnas.0914142107
Lens epithelium-derived growth factor fusion proteins redirect HIV-1 DNA integration
Abstract
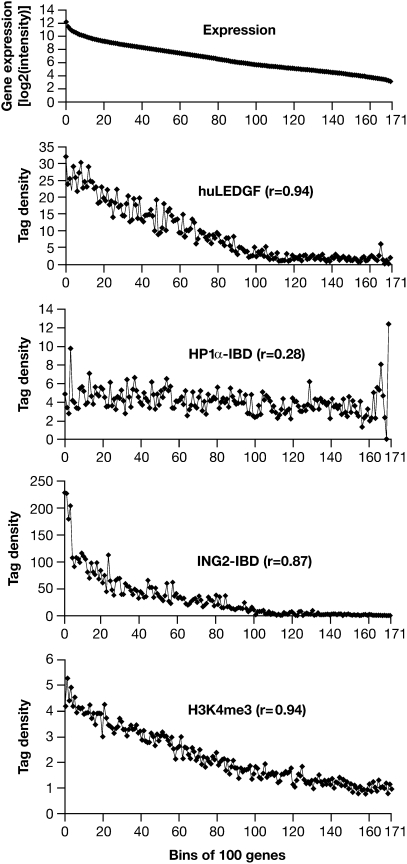
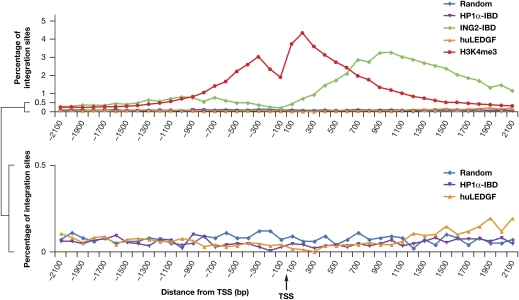
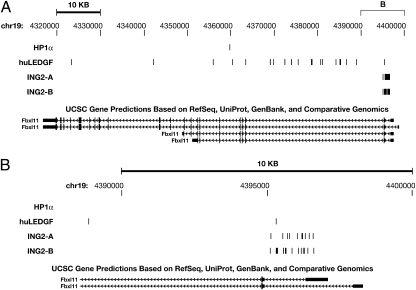
Lens epithelium-derived growth factor (LEDGF) fusion proteins can direct HIV-1 DNA integration to novel sites in the host genome. The C terminus of LEDGF contains an integrase binding domain (IBD), and the N terminus binds chromatin. LEDGF normally directs integrations to the bodies of expressed genes. Replacing the N terminus of LEDGF with chromatin binding domains (CBDs) from other proteins changes the specificity of HIV-1 DNA integration. We chose two well-characterized CBDs: the plant homeodomain (PHD) finger from ING2 and the chromodomain from heterochromatin binding protein 1alpha (HP1alpha). The ING2 PHD finger binds H3K4me3, a histone mark that is associated with the transcriptional start sites of expressed genes. The HP1alpha chromodomain binds H3K9me2,3, histone marks that are widely distributed throughout the genome. A fusion protein in which the ING2 PHD finger was linked to the LEDGF IBD directed integrations near the start sites of expressed genes. A similar fusion protein in which the HP1alpha chromodomain was linked to the LEDGF IBD directed integrations to sites that differed from both the PHD finger fusion-directed and LEDGF-directed integration sites. The ability to redirect HIV-1 DNA integration may help solve the problems associated with the activation of oncogenes when retroviruses are used in gene therapy.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Comment in
-
Targeting HIV-1 DNA integration by swapping tethers.Proc Natl Acad Sci U S A. 2010 Feb 16;107(7):2735-6. doi: 10.1073/pnas.0915097107. Epub 2010 Feb 9. Proc Natl Acad Sci U S A. 2010. PMID: 20145107 Free PMC article. No abstract available.
Similar articles
-
Implication of serine residues 271, 273, and 275 in the human immunodeficiency virus type 1 cofactor activity of lens epithelium-derived growth factor/p75.J Virol. 2010 Jan;84(2):740-52. doi: 10.1128/JVI.01043-09. Epub 2009 Nov 4. J Virol. 2010. PMID: 19889764 Free PMC article.
-
Characterization of the HIV-1 integrase chromatin- and LEDGF/p75-binding abilities by mutagenic analysis within the catalytic core domain of integrase.Virol J. 2010 Mar 23;7:68. doi: 10.1186/1743-422X-7-68. Virol J. 2010. PMID: 20331877 Free PMC article.
-
The Structure-Specific Recognition Protein 1 Associates with Lens Epithelium-Derived Growth Factor Proteins and Modulates HIV-1 Replication.J Mol Biol. 2016 Jul 17;428(14):2814-31. doi: 10.1016/j.jmb.2016.05.013. Epub 2016 May 21. J Mol Biol. 2016. PMID: 27216501 Free PMC article.
-
Cellular co-factors of HIV-1 integration.Trends Biochem Sci. 2006 Feb;31(2):98-105. doi: 10.1016/j.tibs.2005.12.002. Epub 2006 Jan 5. Trends Biochem Sci. 2006. PMID: 16403635 Review.
-
Inhibitors of the interactions between HIV-1 IN and the cofactor LEDGF/p75.ChemMedChem. 2011 Jul 4;6(7):1184-91. doi: 10.1002/cmdc.201100071. Epub 2011 Apr 19. ChemMedChem. 2011. PMID: 21506277 Review.
Cited by
-
Strategies for Targeting Retroviral Integration for Safer Gene Therapy: Advances and Challenges.Front Mol Biosci. 2021 May 12;8:662331. doi: 10.3389/fmolb.2021.662331. eCollection 2021. Front Mol Biosci. 2021. PMID: 34055882 Free PMC article. Review.
-
Lens epithelium-derived growth factor/p75 qualifies as a target for HIV gene therapy in the NSG mouse model.Mol Ther. 2012 May;20(5):908-17. doi: 10.1038/mt.2012.6. Epub 2012 Feb 14. Mol Ther. 2012. PMID: 22334021 Free PMC article.
-
Cellular and molecular mechanisms of HIV-1 integration targeting.Cell Mol Life Sci. 2018 Jul;75(14):2491-2507. doi: 10.1007/s00018-018-2772-5. Epub 2018 Feb 7. Cell Mol Life Sci. 2018. PMID: 29417178 Free PMC article. Review.
-
An analytical pipeline for identifying and mapping the integration sites of HIV and other retroviruses.BMC Genomics. 2020 Mar 9;21(1):216. doi: 10.1186/s12864-020-6647-4. BMC Genomics. 2020. PMID: 32151239 Free PMC article.
-
Allosteric inhibition of human immunodeficiency virus integrase: late block during viral replication and abnormal multimerization involving specific protein domains.J Biol Chem. 2014 Jul 25;289(30):20477-88. doi: 10.1074/jbc.M114.551119. J Biol Chem. 2014. PMID: 24904063 Free PMC article.
References
-
- Hughes SH, et al. Proviruses of avian sarcoma virus are terminally redundant, co-extensive with unintegrated linear DNA and integrated at many sites. Cell. 1978;15:1397–1410. - PubMed
-
- Wu X, Li Y, Crise B, Burgess SM. Transcription start regions in the human genome are favored targets for MLV integration. Science. 2003;300:1749–1751. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
