

Epstein-Barr virus (EBV)-encoded small RNA is released from EBV-infected cells and activates signaling from Toll-like receptor 3
- PMID: 19720839
- PMCID: PMC2757889
- DOI: 10.1084/jem.20081761
Epstein-Barr virus (EBV)-encoded small RNA is released from EBV-infected cells and activates signaling from Toll-like receptor 3
Abstract
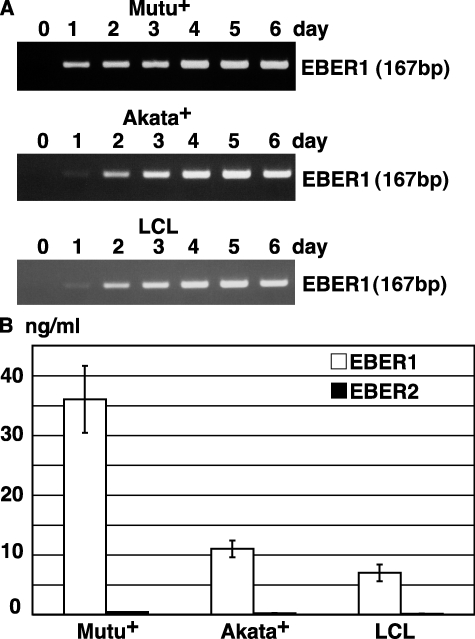
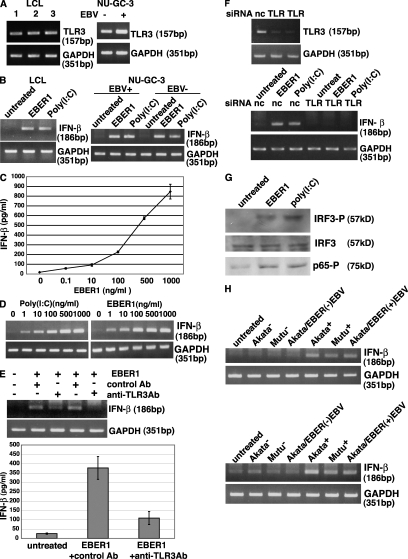
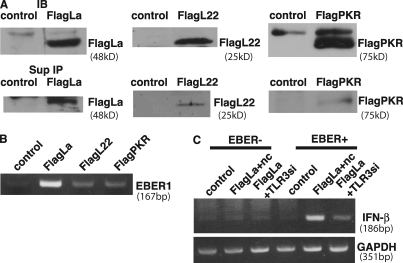
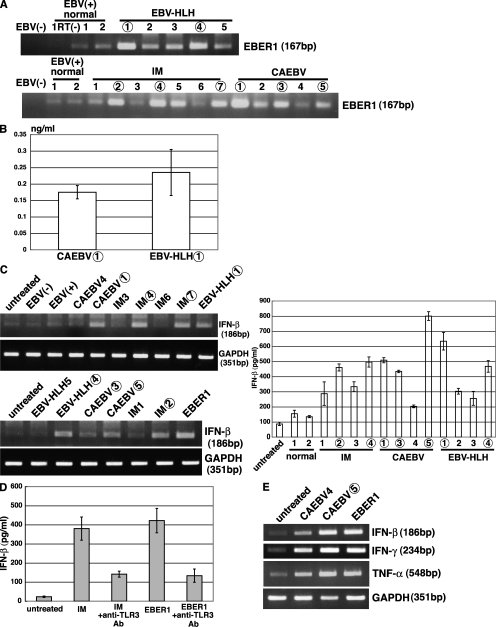
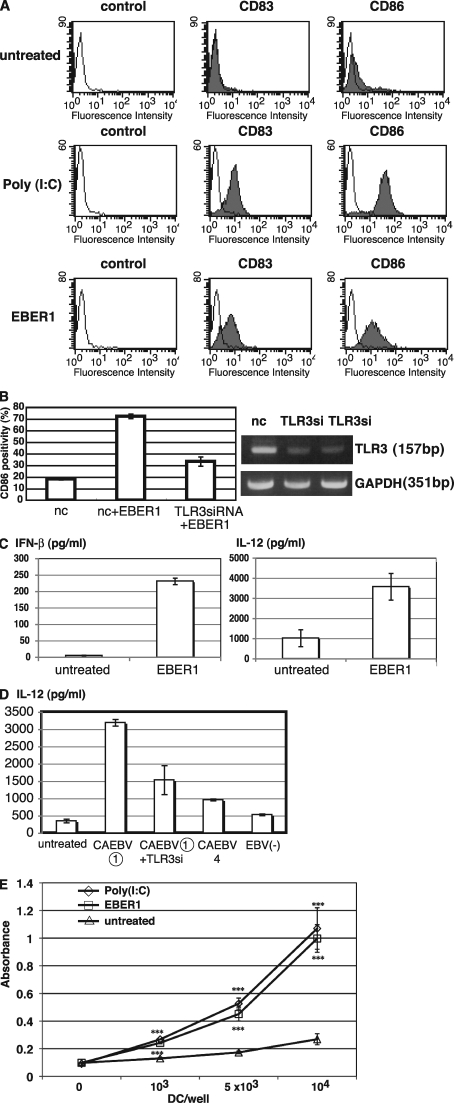
Epstein-Barr virus-encoded small RNA (EBER) is nonpolyadenylated, noncoding RNA that forms stem-loop structure by intermolecular base-pairing, giving rise to double-stranded RNA (dsRNA)-like molecules, and exists abundantly in EBV-infected cells. Here, we report that EBER induces signaling from the Toll-like receptor 3 (TLR3), which is a sensor of viral double-stranded RNA (dsRNA) and induces type I IFN and proinflammatory cytokines. A substantial amount of EBER, which was sufficient to induce signaling from TLR3, was released from EBV-infected cells, and the majority of the released EBER existed as a complex with a cellular EBER-binding protein La, suggesting that EBER was released from the cells by active secretion of La. Sera from patients with infectious mononucleosis (IM), chronic active EBV infection (CAEBV), and EBV-associated hemophagocytic lymphohistiocytosis (EBV-HLH), whose general symptoms are caused by proinflammatory cytokines contained EBER, and addition of RNA purified from the sera into culture medium induced signaling from TLR3 in EBV-transformed lymphocytes and peripheral mononuclear cells. Furthermore, DCs treated with EBER showed mature phenotype and antigen presentation capacity. These findings suggest that EBER, which is released from EBV-infected cells, is responsible for immune activation by EBV, inducing type I IFN and proinflammatory cytokines. EBER-induced activation of innate immunity would account for immunopathologic diseases caused by active EBV infection.
Figures
Similar articles
-
Epstein-Barr virus-encoded small RNA induces IL-10 through RIG-I-mediated IRF-3 signaling.Oncogene. 2008 Jul 10;27(30):4150-60. doi: 10.1038/onc.2008.75. Epub 2008 Mar 24. Oncogene. 2008. PMID: 18362887
-
Epstein-Barr virus-encoded small RNAs (EBERs) are present in fractions related to exosomes released by EBV-transformed cells.PLoS One. 2014 Jun 4;9(6):e99163. doi: 10.1371/journal.pone.0099163. eCollection 2014. PLoS One. 2014. PMID: 24896633 Free PMC article.
-
[Molecular mechanisms of Epstein-Barr virus-mediated carcinogeneis].Uirusu. 2014;64(1):49-56. doi: 10.2222/jsv.64.49. Uirusu. 2014. PMID: 25765980 Review. Japanese.
-
Epstein-Barr Virus-Encoded RNAs: Key Molecules in Viral Pathogenesis.Cancers (Basel). 2014 Aug 6;6(3):1615-30. doi: 10.3390/cancers6031615. Cancers (Basel). 2014. PMID: 25101570 Free PMC article. Review.
-
EBV-encoded RNA via TLR3 induces inflammation in nasopharyngeal carcinoma.Oncotarget. 2015 Sep 15;6(27):24291-303. doi: 10.18632/oncotarget.4552. Oncotarget. 2015. PMID: 26172457 Free PMC article.
Cited by
-
Significance and Implications of Patient-reported Xerostomia in Sjögren's Syndrome: Findings From the National Institutes of Health Cohort.EBioMedicine. 2016 Oct;12:270-279. doi: 10.1016/j.ebiom.2016.09.005. Epub 2016 Sep 9. EBioMedicine. 2016. PMID: 27639822 Free PMC article.
-
Viral non-coding RNAs: Stealth strategies in the tug-of-war between humans and herpesviruses.Semin Cell Dev Biol. 2021 Mar;111:135-147. doi: 10.1016/j.semcdb.2020.06.015. Epub 2020 Jul 3. Semin Cell Dev Biol. 2021. PMID: 32631785 Free PMC article. Review.
-
Trial watch: TLR3 agonists in cancer therapy.Oncoimmunology. 2020 Jun 2;9(1):1771143. doi: 10.1080/2162402X.2020.1771143. Oncoimmunology. 2020. PMID: 32934877 Free PMC article. Review.
-
Innate immune responses against Epstein Barr virus infection.J Leukoc Biol. 2013 Dec;94(6):1185-90. doi: 10.1189/jlb.0313173. Epub 2013 Jun 28. J Leukoc Biol. 2013. PMID: 23812328 Free PMC article. Review.
-
Latency type-dependent modulation of Epstein-Barr virus-encoded latent membrane protein 1 expression by type I interferons in B cells.J Virol. 2012 Apr;86(8):4701-7. doi: 10.1128/JVI.06829-11. Epub 2012 Feb 15. J Virol. 2012. PMID: 22345482 Free PMC article.
References
-
- Akira S., Takeda K. 2004. Toll-like receptor signalling.Nat. Rev. Immunol. 4:499–511 doi:10.1038/nri1391 - DOI - PubMed
-
- Akiyama S., Amo H., Watanabe T., Matsuyama M., Sakamoto J., Imaizumi M., Ichihashi H., Kondo T., Takagi H. 1988. Characteristics of three human gastric cancer cell lines, NU-GC-2, NU-GC-3 and NU-GC-4.Jpn. J. Surg. 18:438–446 doi:10.1007/BF02471470 - DOI - PubMed
-
- Clarke P.A., Schwemmle M., Schickinger J., Hilse K., Clemens M.J. 1991. Binding of Epstein-Barr virus small RNA EBER-1 to the double-stranded RNA-activated protein kinase DAI.Nucleic Acids Res. 19:243–248 doi:10.1093/nar/19.2.243 - DOI - PMC - PubMed
-
- Clarke P.A., Sharp N.A., Clemens M.J. 1992. Expression of genes for the Epstein-Barr virus small RNAs EBER-1 and EBER-2 in Daudi Burkitt's lymphoma cells: effects of interferon treatment.J. Gen. Virol. 73:3169–3175 doi:10.1099/0022-1317-73-12-3169 - DOI - PubMed
-
- Doyle S.E., O'Connell R., Vaidya S.A., Chow E.K., Yee K., Cheng G. 2003. Toll-like receptor 3 mediates a more potent antiviral response than Toll-like receptor 4.J. Immunol. 170:3565–3571 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
