

Mechanisms that regulate localization of a DNA double-strand break to the nuclear periphery
- PMID: 19390086
- PMCID: PMC2675867
- DOI: 10.1101/gad.1782209
Mechanisms that regulate localization of a DNA double-strand break to the nuclear periphery
Abstract
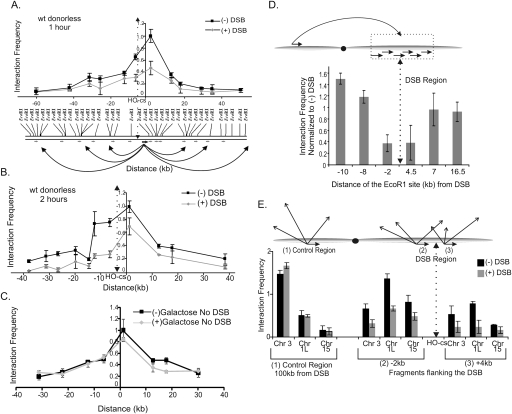
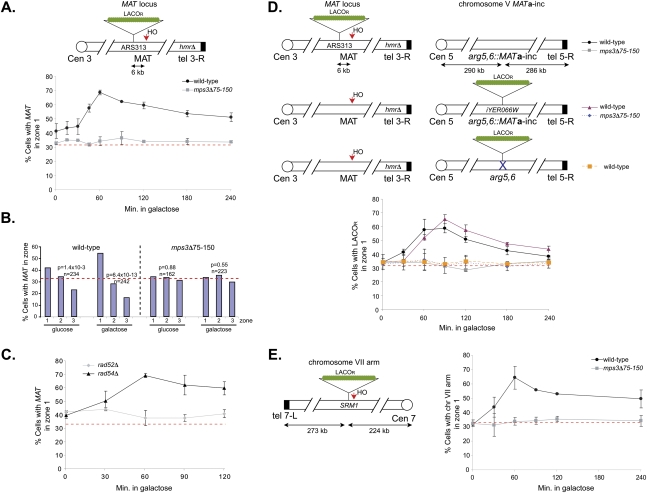
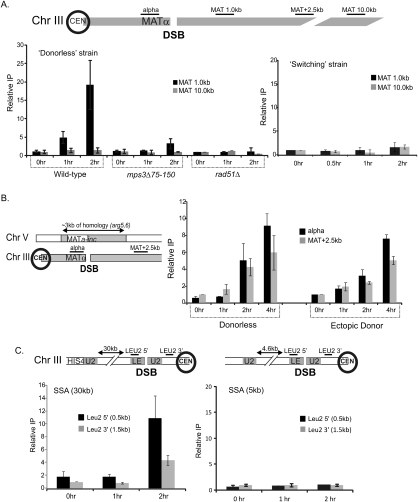
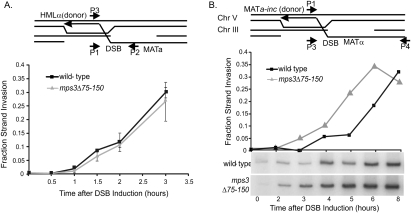
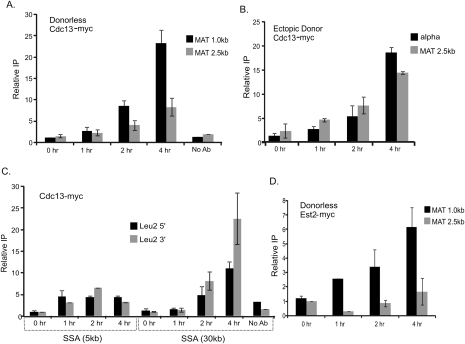
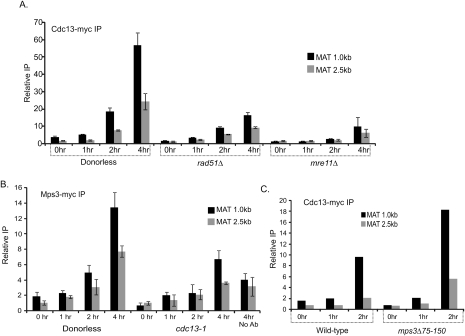
DNA double-strand breaks (DSBs) are among the most deleterious forms of DNA lesions in cells. Here we induced site-specific DSBs in yeast cells and monitored chromatin dynamics surrounding the DSB using Chromosome Conformation Capture (3C). We find that formation of a DSB within G1 cells is not sufficient to alter chromosome dynamics. However, DSBs formed within an asynchronous cell population result in large decreases in both intra- and interchromosomal interactions. Using live cell microscopy, we find that changes in chromosome dynamics correlate with relocalization of the DSB to the nuclear periphery. Sequestration to the periphery requires the nuclear envelope protein, Mps3p, and Mps3p-dependent tethering delays recombinational repair of a DSB and enhances gross chromosomal rearrangements. Furthermore, we show that components of the telomerase machinery are recruited to a DSB and that telomerase recruitment is required for its peripheral localization. Based on these findings, we propose that sequestration of unrepaired or slowly repaired DSBs to the nuclear periphery reflects a competition between alternative repair pathways.
Figures

Similar articles
-
Asymmetric Processing of DNA Ends at a Double-Strand Break Leads to Unconstrained Dynamics and Ectopic Translocation.Cell Rep. 2018 Sep 4;24(10):2614-2628.e4. doi: 10.1016/j.celrep.2018.07.102. Cell Rep. 2018. PMID: 30184497
-
Telomere tethering at the nuclear periphery is essential for efficient DNA double strand break repair in subtelomeric region.J Cell Biol. 2006 Jan 16;172(2):189-99. doi: 10.1083/jcb.200505159. J Cell Biol. 2006. PMID: 16418532 Free PMC article.
-
Cell cycle-dependent spatial segregation of telomerase from sites of DNA damage.J Cell Biol. 2017 Aug 7;216(8):2355-2371. doi: 10.1083/jcb.201610071. Epub 2017 Jun 21. J Cell Biol. 2017. PMID: 28637749 Free PMC article.
-
Chromosome healing by de novo telomere addition in Saccharomyces cerevisiae.Mol Microbiol. 2006 Mar;59(5):1357-68. doi: 10.1111/j.1365-2958.2006.05026.x. Mol Microbiol. 2006. PMID: 16468981 Review.
-
Emerging non-canonical roles for the Rad51-Rad52 interaction in response to double-strand breaks in yeast.Curr Genet. 2020 Oct;66(5):917-926. doi: 10.1007/s00294-020-01081-z. Epub 2020 May 12. Curr Genet. 2020. PMID: 32399607 Free PMC article. Review.
Cited by
-
The SUN protein Mps3 is required for spindle pole body insertion into the nuclear membrane and nuclear envelope homeostasis.PLoS Genet. 2011 Nov;7(11):e1002365. doi: 10.1371/journal.pgen.1002365. Epub 2011 Nov 17. PLoS Genet. 2011. PMID: 22125491 Free PMC article.
-
Nuclear envelope tethering inhibits the formation of ALT-associated PML bodies in ALT cells.Aging (Albany NY). 2021 Apr 4;13(7):10490-10516. doi: 10.18632/aging.202810. Epub 2021 Apr 4. Aging (Albany NY). 2021. PMID: 33820871 Free PMC article.
-
Nuclear Cytoskeleton in Virus Infection.Int J Mol Sci. 2022 Jan 5;23(1):578. doi: 10.3390/ijms23010578. Int J Mol Sci. 2022. PMID: 35009004 Free PMC article. Review.
-
DNA-end capping by the budding yeast transcription factor and subtelomeric binding protein Tbf1.EMBO J. 2012 Jan 4;31(1):138-49. doi: 10.1038/emboj.2011.349. Epub 2011 Sep 27. EMBO J. 2012. PMID: 21952045 Free PMC article.
-
Interchromosomal homology searches drive directional ALT telomere movement and synapsis.Cell. 2014 Sep 25;159(1):108-121. doi: 10.1016/j.cell.2014.08.030. Cell. 2014. PMID: 25259924 Free PMC article.
References
-
- Antoniacci L.M., Kenna M.A., Uetz P., Fields S., Skibbens R.V. The spindle pole body assembly component Mps3p/Nep98p functions in sister chromatid cohesion. J. Biol. Chem. 2004;279:49542–49550. - PubMed
-
- Antoniacci L.M., Kenna M.A., Skibbens R.V. The nuclear envelope and spindle pole body-associated Mps3 protein bind telomere regulators and function in telomere clustering. Cell Cycle. 2007;6:75–79. - PubMed
-
- Aten J.A., Stap J., Krawczyk P.M., van Oven C.H., Hoebe R.A., Essers J., Kanaar R. Dynamics of DNA double-strand breaks revealed by clustering of damaged chromosome domains. Science. 2004;303:92–95. - PubMed
-
- Bakkenist C.J., Kastan M.B. DNA damage activates ATM through intermolecular autophosphorylation and dimer dissociation. Nature. 2003;421:499–506. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases