

Autophagy is an essential component of Drosophila immunity against vesicular stomatitis virus
- PMID: 19362021
- PMCID: PMC2754303
- DOI: 10.1016/j.immuni.2009.02.009
Autophagy is an essential component of Drosophila immunity against vesicular stomatitis virus
Abstract
Intrinsic innate immune mechanisms are the first line of defense against pathogens and exist to control infection autonomously in infected cells. Here, we showed that autophagy, an intrinsic mechanism that can degrade cytoplasmic components, played a direct antiviral role against the mammalian viral pathogen vesicular stomatitis virus (VSV) in the model organism Drosophila. We found that the surface glycoprotein, VSV-G, was likely the pathogen-associated molecular pattern (PAMP) that initiated this cell-autonomous response. Once activated, autophagy decreased viral replication, and repression of autophagy led to increased viral replication and pathogenesis in cells and animals. Lastly, we showed that the antiviral response was controlled by the phosphatidylinositol 3-kinase (PI3K)-Akt-signaling pathway, which normally regulates autophagy in response to nutrient availability. Altogether, these data uncover an intrinsic antiviral program that links viral recognition to the evolutionarily conserved nutrient-signaling and autophagy pathways.
Figures
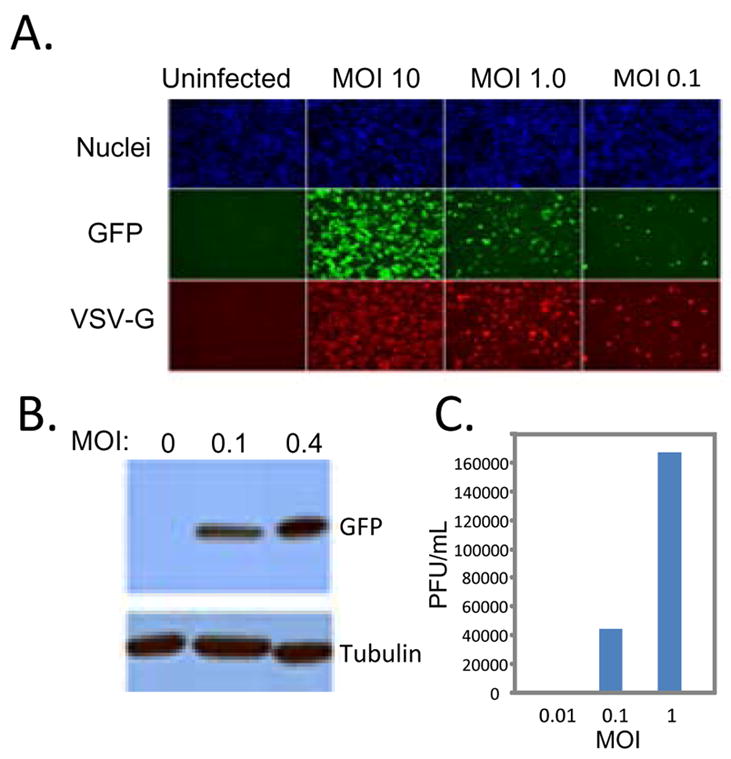
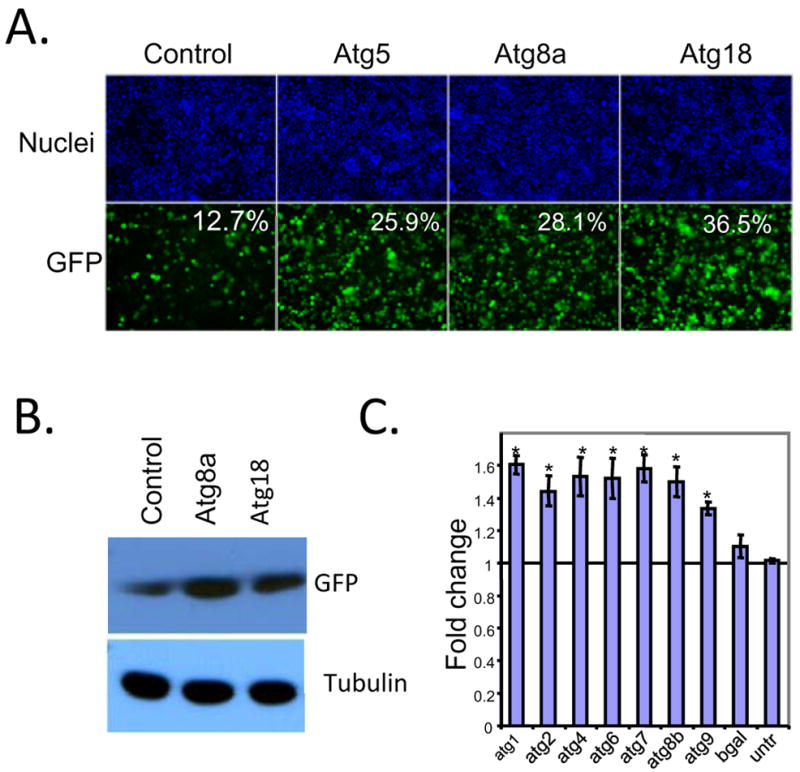
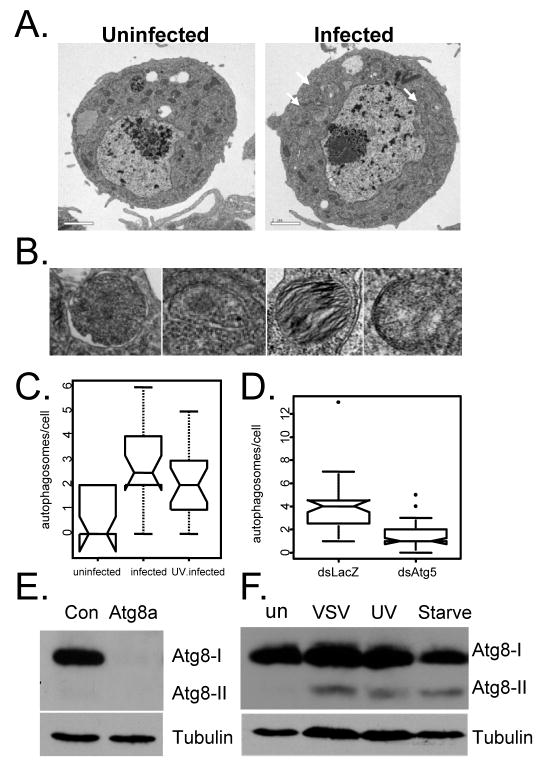
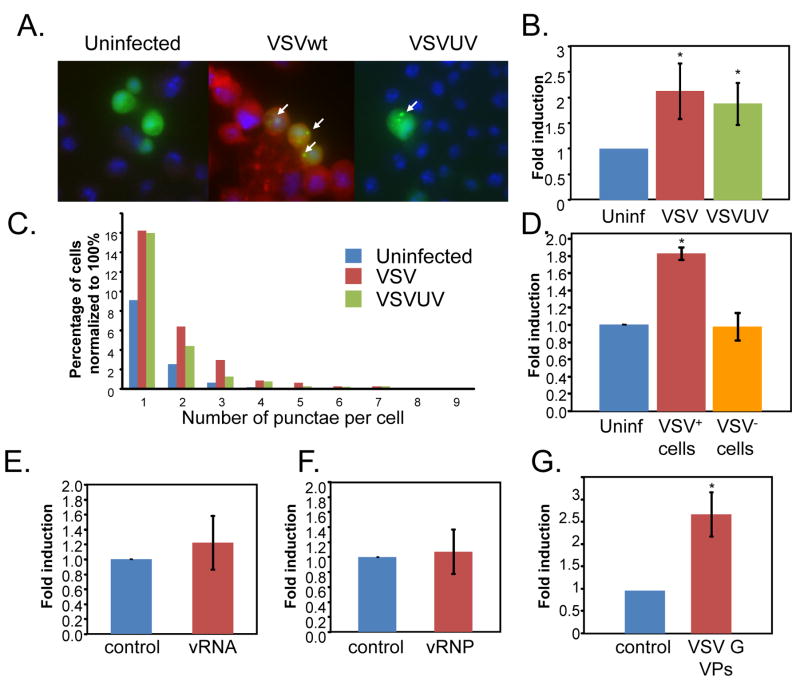
Comment in
-
Strange bedfellows expose ancient secrets of autophagy in immunity.Immunity. 2009 Apr 17;30(4):479-81. doi: 10.1016/j.immuni.2009.03.005. Immunity. 2009. PMID: 19371713 Free PMC article.
Similar articles
-
VSV infection is sensed by Drosophila, attenuates nutrient signaling, and thereby activates antiviral autophagy.Autophagy. 2009 Oct;5(7):1062-3. doi: 10.4161/auto.5.7.9730. Epub 2009 Oct 5. Autophagy. 2009. PMID: 19713743
-
Analysis of the Contribution of Hemocytes and Autophagy to Drosophila Antiviral Immunity.J Virol. 2016 May 12;90(11):5415-5426. doi: 10.1128/JVI.00238-16. Print 2016 Jun 1. J Virol. 2016. PMID: 27009948 Free PMC article.
-
Virus recognition by Toll-7 activates antiviral autophagy in Drosophila.Immunity. 2012 Apr 20;36(4):658-67. doi: 10.1016/j.immuni.2012.03.003. Epub 2012 Mar 29. Immunity. 2012. PMID: 22464169 Free PMC article.
-
Autophagy and innate immunity: Insights from invertebrate model organisms.Autophagy. 2018;14(2):233-242. doi: 10.1080/15548627.2017.1389824. Epub 2018 Feb 17. Autophagy. 2018. PMID: 29130360 Free PMC article. Review.
-
Drosophila immunity against natural and nonnatural viral pathogens.Virology. 2020 Jan 15;540:165-171. doi: 10.1016/j.virol.2019.12.001. Epub 2019 Dec 1. Virology. 2020. PMID: 31928998 Review.
Cited by
-
Transcriptional pausing controls a rapid antiviral innate immune response in Drosophila.Cell Host Microbe. 2012 Oct 18;12(4):531-43. doi: 10.1016/j.chom.2012.08.011. Cell Host Microbe. 2012. PMID: 23084920 Free PMC article.
-
Programmed autophagy in the fat body of Aedes aegypti is required to maintain egg maturation cycles.PLoS One. 2011;6(11):e25502. doi: 10.1371/journal.pone.0025502. Epub 2011 Nov 17. PLoS One. 2011. PMID: 22125592 Free PMC article.
-
Baculovirus infection triggers a shift from amino acid starvation-induced autophagy to apoptosis.PLoS One. 2012;7(5):e37457. doi: 10.1371/journal.pone.0037457. Epub 2012 May 21. PLoS One. 2012. PMID: 22629397 Free PMC article.
-
HijAkt: The PI3K/Akt pathway in virus replication and pathogenesis.Prog Mol Biol Transl Sci. 2012;106:223-50. doi: 10.1016/B978-0-12-396456-4.00002-X. Prog Mol Biol Transl Sci. 2012. PMID: 22340720 Free PMC article. Review.
-
The effects of autophagy on the replication of Nelson Bay orthoreovirus.Virol J. 2019 Jul 18;16(1):90. doi: 10.1186/s12985-019-1196-7. Virol J. 2019. PMID: 31319897 Free PMC article.
References
-
- Arico S, Petiot A, Bauvy C, Dubbelhuis PF, Meijer AJ, Codogno P, Ogier-Denis E. The tumor suppressor PTEN positively regulates macroautophagy by inhibiting the phosphatidylinositol 3-kinase/protein kinase B pathway. J Biol Chem. 2001;276:35243–35246. - PubMed
-
- Brand AH, Perrimon N. Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development. 1993;118:401–415. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
