

PICK1 deficiency causes male infertility in mice by disrupting acrosome formation
- PMID: 19258705
- PMCID: PMC2662547
- DOI: 10.1172/JCI36230
PICK1 deficiency causes male infertility in mice by disrupting acrosome formation
Abstract
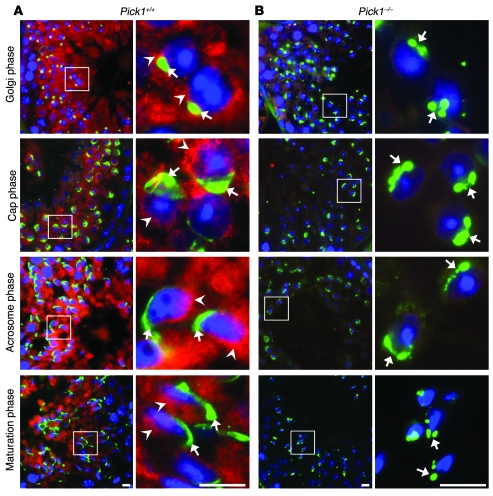
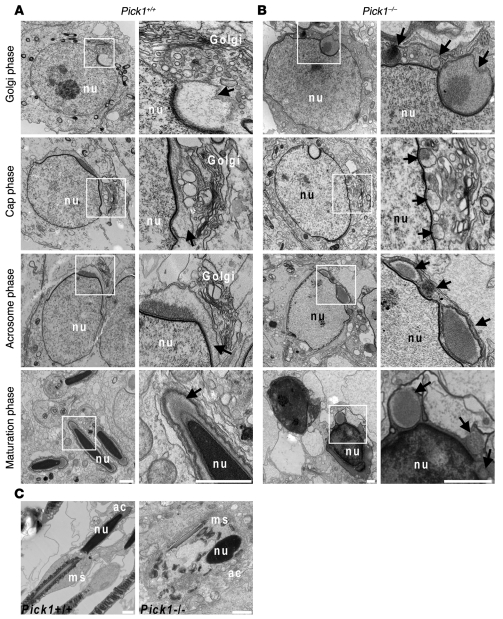
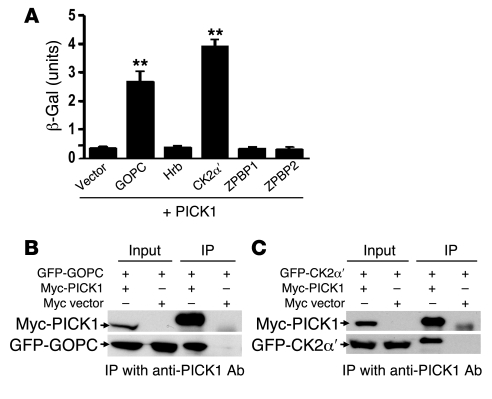
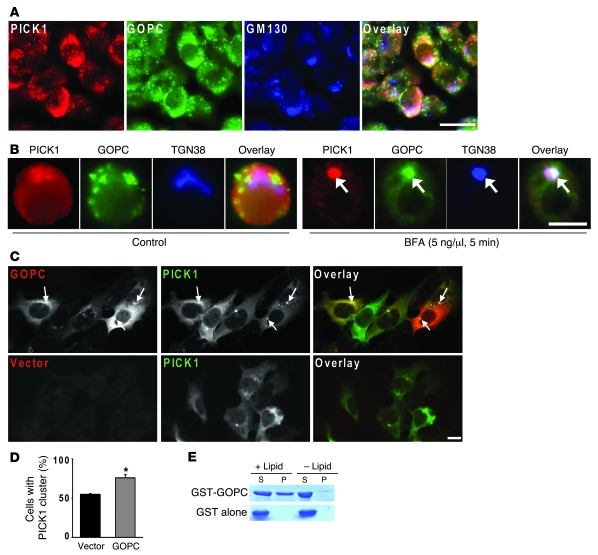
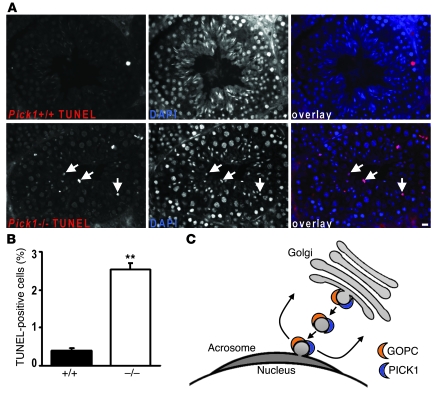
Protein interacting with C kinase 1 (PICK1) is a peripheral membrane protein involved in protein trafficking, a function that has been well characterized in neurons. Here, we report that male mice deficient in PICK1 are infertile and have a phenotype resembling the human disease globozoospermia. The primary defect in the testes of Pick1-knockout mice was fragmentation of acrosomes in the early stages of spermiogenesis. This fragmentation was followed by defects in nuclear elongation and mitochondrial sheath formation, leading to round-headed sperm, reduced sperm count, and severely impaired sperm motility. We found that PICK1 interacted with Golgi-associated PDZ- and coiled-coil motif-containing protein (GOPC) and the primary catalytic subunit of protein kinase 2 (CK2alpha'), proteins whose deficiencies lead to globozoospermia in mice. PICK1 was highly expressed in round spermatids and localized to Golgi-derived proacrosomal granules. GOPC colocalized with PICK1 in the Golgi region and facilitated formation of PICK1-positive clusters. Furthermore, there was an increase in apoptosis in the seminiferous tubules of Pick1-/- mice, a phenotype also seen in CK2alpha'-deficient mice. Our results suggest that PICK1 is involved in vesicle trafficking from the Golgi apparatus to the acrosome and cooperates with other proteins such as GOPC and CK2alpha' in acrosome biogenesis.
Figures



Similar articles
-
ICA1L forms BAR-domain complexes with PICK1 and is crucial for acrosome formation in spermiogenesis.J Cell Sci. 2015 Oct 15;128(20):3822-36. doi: 10.1242/jcs.173534. Epub 2015 Aug 25. J Cell Sci. 2015. PMID: 26306493
-
Failure to assemble the peri-nuclear structures in GOPC deficient spermatids as found in round-headed spermatozoa.Arch Histol Cytol. 2004 Nov;67(4):349-60. doi: 10.1679/aohc.67.349. Arch Histol Cytol. 2004. PMID: 15700542
-
Sirt1 regulates acrosome biogenesis by modulating autophagic flux during spermiogenesis in mice.Development. 2017 Feb 1;144(3):441-451. doi: 10.1242/dev.147074. Epub 2016 Dec 21. Development. 2017. PMID: 28003215
-
Molecular cytogenetic and genetic aspects of globozoospermia: a review.Andrologia. 2013 Feb;45(1):1-9. doi: 10.1111/j.1439-0272.2012.01308.x. Epub 2012 May 10. Andrologia. 2013. PMID: 22571172 Review.
-
[Sperm acrosome formation-associated genes in mice: Advances in studies].Zhonghua Nan Ke Xue. 2016 Jan;22(1):72-6. Zhonghua Nan Ke Xue. 2016. PMID: 26931031 Review. Chinese.
Cited by
-
Decreased Expression of KIFC1 in Human Testes with Globozoospermic Defects.Genes (Basel). 2016 Sep 27;7(10):75. doi: 10.3390/genes7100075. Genes (Basel). 2016. PMID: 27690105 Free PMC article.
-
An Overview of The Globozoospermia as A Multigenic Identified Syndrome.Int J Fertil Steril. 2019 Jan;12(4):273-277. doi: 10.22074/ijfs.2019.5561. Epub 2018 Oct 2. Int J Fertil Steril. 2019. PMID: 30291685 Free PMC article.
-
PICK1 regulates the trafficking of ASIC1a and acidotoxicity in a BAR domain lipid binding-dependent manner.Mol Brain. 2010 Dec 21;3:39. doi: 10.1186/1756-6606-3-39. Mol Brain. 2010. PMID: 21176140 Free PMC article.
-
The Arf GAP SMAP2 is necessary for organized vesicle budding from the trans-Golgi network and subsequent acrosome formation in spermiogenesis.Mol Biol Cell. 2013 Sep;24(17):2633-44. doi: 10.1091/mbc.E13-05-0234. Epub 2013 Jul 17. Mol Biol Cell. 2013. PMID: 23864717 Free PMC article.
-
TBC1D20 mediates autophagy as a key regulator of autophagosome maturation.Autophagy. 2016 Oct 2;12(10):1759-1775. doi: 10.1080/15548627.2016.1199300. Epub 2016 Aug 3. Autophagy. 2016. PMID: 27487390 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
