

Nuclear sphingolipids: metabolism and signaling
- PMID: 18326892
- PMCID: PMC2386903
- DOI: 10.1194/jlr.R800009-JLR200
Nuclear sphingolipids: metabolism and signaling
Abstract
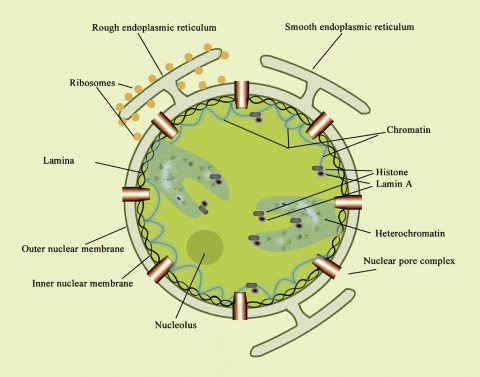
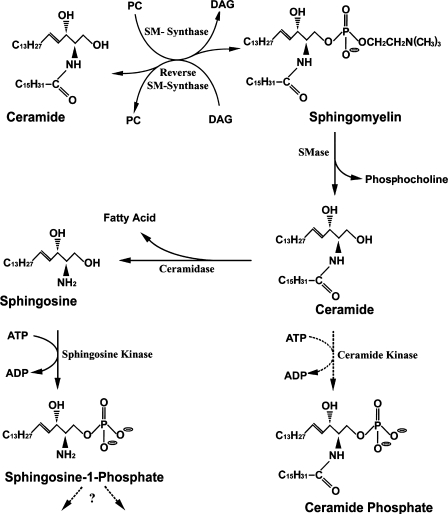
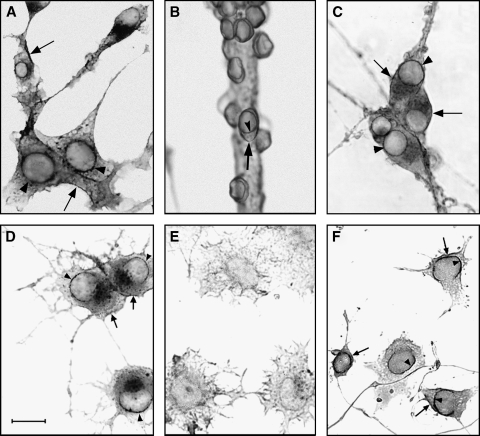
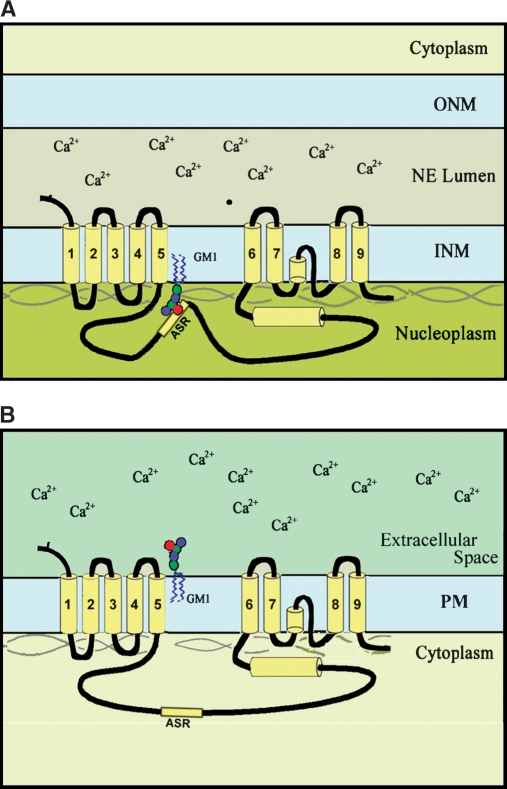
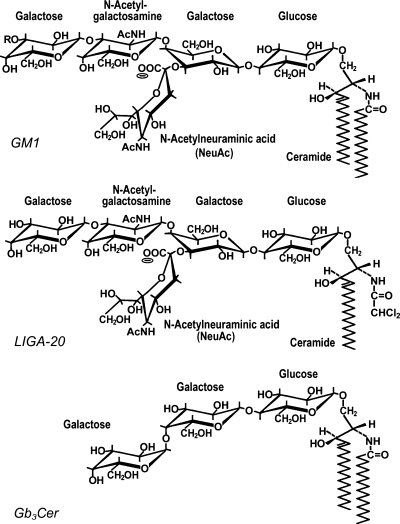
Sphingolipids are most prominently expressed in the plasma membrane, but recent studies have pointed to important signaling and regulatory roles in the nucleus. The most abundant nuclear sphingolipid is sphingomyelin (SM), which occurs in the nuclear envelope (NE) as well as intranuclear sites. The major metabolic product of SM is ceramide, which is generated by nuclear sphingomyelinase and triggers apoptosis and other metabolic changes. Ceramide is further hydrolyzed to free fatty acid and sphingosine, the latter undergoing conversion to sphingosine phosphate by action of a specific nuclear kinase. Gangliosides are another type of sphingolipid found in the nucleus, members of the a-series of gangliotetraose gangliosides (GM1, GD1a) occurring in the NE and endonuclear compartments. GM1 in the inner membrane of the NE is tightly associated with a Na(+)/Ca(2+) exchanger whose activity it potentiates, thereby contributing to regulation of Ca(2+) homeostasis in the nucleus. This was shown to exert a cytoprotective role as absence or inactivation of this nuclear complex rendered cells vulnerable to apoptosis. This was demonstrated in the greatly enhanced kainite-induced seizure activity in knockout mice lacking gangliotetraose gangliosides. The pathology included apoptotic destruction of neurons in the CA3 region of the hippocampus. Ca(2+) homeostasis was restored in these animals with LIGA-20, a membrane-permeant derivative of GM1 that entered the NE and activated the nuclear Na(+)/Ca(2+) exchanger. Some evidence suggests the presence of uncharged glycosphingolipids in the nucleus.
Figures
Similar articles
-
Sphingolipids of the nucleus and their role in nuclear signaling.Biochim Biophys Acta. 2006 May-Jun;1761(5-6):588-98. doi: 10.1016/j.bbalip.2006.04.010. Epub 2006 May 6. Biochim Biophys Acta. 2006. PMID: 16814200 Review.
-
Gangliosides of the nuclear membrane: a crucial locus of cytoprotective modulation.J Cell Biochem. 2006 Apr 1;97(5):893-903. doi: 10.1002/jcb.20731. J Cell Biochem. 2006. PMID: 16408301 Review.
-
New findings on nuclear gangliosides: overview on metabolism and function.J Neurochem. 2011 Mar;116(5):714-20. doi: 10.1111/j.1471-4159.2010.07115.x. Epub 2011 Jan 13. J Neurochem. 2011. PMID: 21214576 Review.
-
Enhanced susceptibility to kainate-induced seizures, neuronal apoptosis, and death in mice lacking gangliotetraose gangliosides: protection with LIGA 20, a membrane-permeant analog of GM1.J Neurosci. 2005 Nov 23;25(47):11014-22. doi: 10.1523/JNEUROSCI.3635-05.2005. J Neurosci. 2005. PMID: 16306414 Free PMC article.
-
Nuclear sphingolipid metabolism.Annu Rev Physiol. 2012;74:131-51. doi: 10.1146/annurev-physiol-020911-153321. Epub 2011 Sep 9. Annu Rev Physiol. 2012. PMID: 21888508 Free PMC article. Review.
Cited by
-
Critical role for the protons in FRTL-5 thyroid cells: nuclear sphingomyelinase induced-damage.Int J Mol Sci. 2014 Jun 27;15(7):11555-65. doi: 10.3390/ijms150711555. Int J Mol Sci. 2014. PMID: 24979136 Free PMC article.
-
Development of Genetically Encoded Fluorescent KSR1-Based Probes to Track Ceramides during Phagocytosis.Int J Mol Sci. 2024 Mar 5;25(5):2996. doi: 10.3390/ijms25052996. Int J Mol Sci. 2024. PMID: 38474242 Free PMC article.
-
Ceramide-induced apoptosis in renal tubular cells: a role of mitochondria and sphingosine-1-phoshate.Int J Mol Sci. 2015 Mar 5;16(3):5076-124. doi: 10.3390/ijms16035076. Int J Mol Sci. 2015. PMID: 25751724 Free PMC article. Review.
-
GM1 asymmetry in the membrane stabilizes pores.Biophys J. 2022 Sep 6;121(17):3295-3302. doi: 10.1016/j.bpj.2022.06.011. Epub 2022 Jun 6. Biophys J. 2022. PMID: 35668647 Free PMC article.
-
The interplay between bioactive sphingolipids and steroid hormones.Steroids. 2010 Jun;75(6):390-9. doi: 10.1016/j.steroids.2010.01.020. Epub 2010 Feb 4. Steroids. 2010. PMID: 20138078 Free PMC article. Review.
References
-
- Chemistry and metabolism of sphingolipids. A collection of papers dedicated to Herbert E. Carter. 1970. Chem. & Phys. Lipids. 5: 1–300.
-
- Goureau M. F., and J. Raulin. 1970. Unsaturation of exogenous fatty acids and composition of phospholipids linked to liver nucleus chromatin. Bull. Soc. Chim. Biol. (Paris). 52 941–953. - PubMed
-
- Cave C., and P. B. Gahan. 1970. A cytochemical and autoradiographic investigation of nuclear phospholipids. Cardiologia. 23 303–312.
-
- Cocco L., N. M. Maraldi, and F. A. Mandzoli. 1980. Phospholipid interactions in rat liver nuclear matrix. Biochem. Biophys. Res. Commun. 9 890–898. - PubMed
-
- Bkaily, G., D. Jacques, and P. D'Orleans-Juste. 2003. Receptors and channels of nuclear membranes as a new target for drug action. In Pathophysiology of Cardiovascular Disease. N. S. Dhalla, H. Rupp, A. Angel, and G. N. Pierce, editors. Kluwer Acad. Publ., Boston. 473–483.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
