

Redox compartmentalization in eukaryotic cells
- PMID: 18267127
- PMCID: PMC2601570
- DOI: 10.1016/j.bbagen.2008.01.011
Redox compartmentalization in eukaryotic cells
Abstract
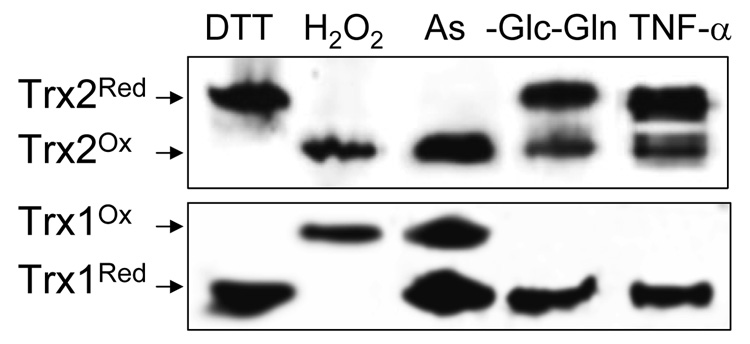
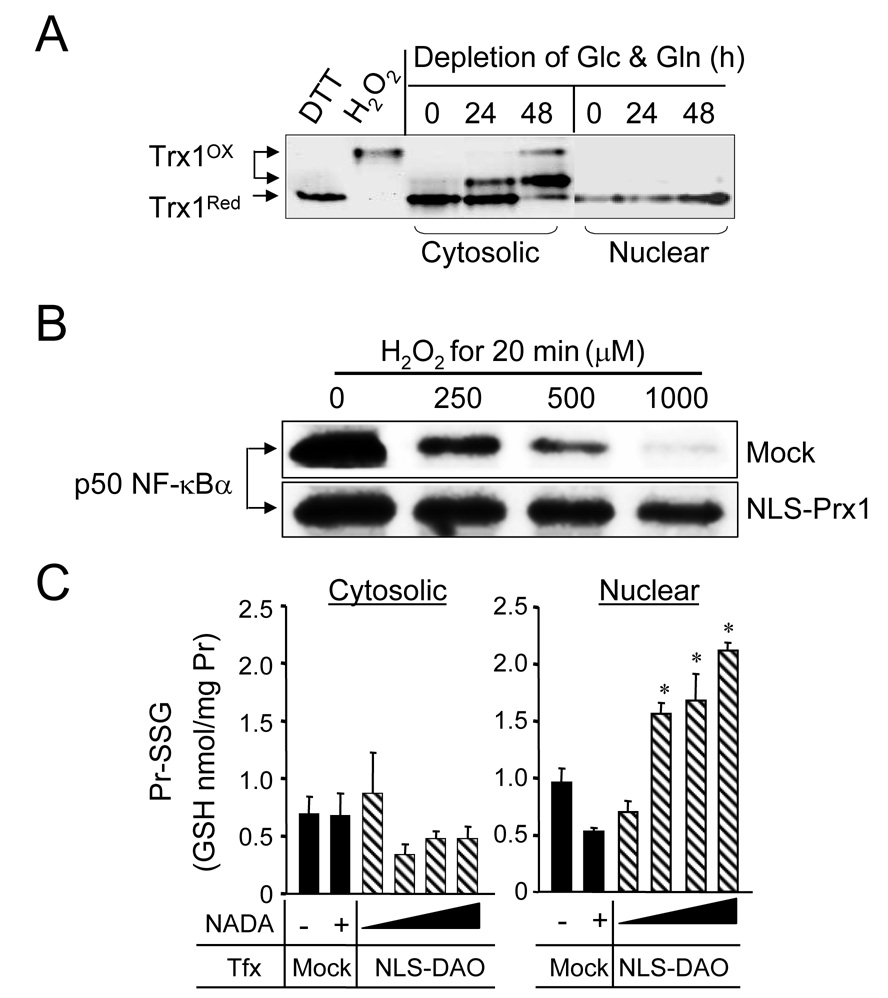
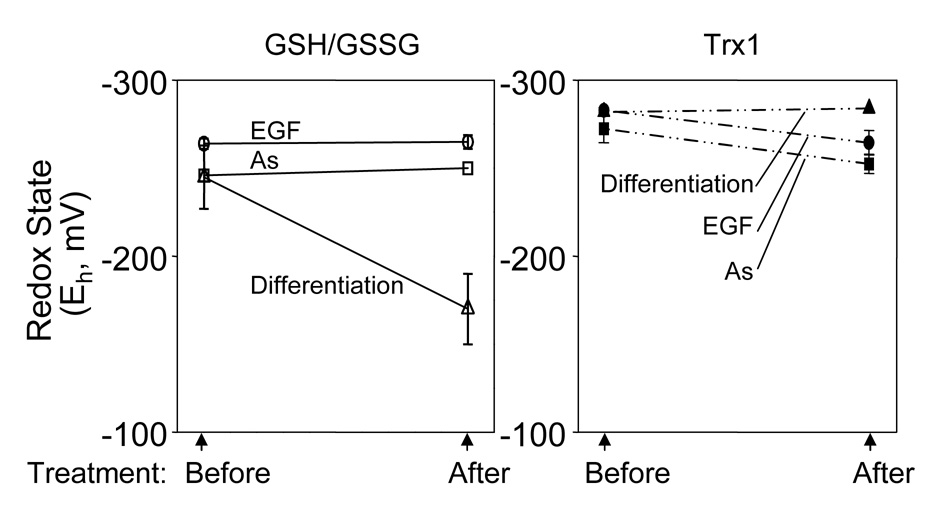
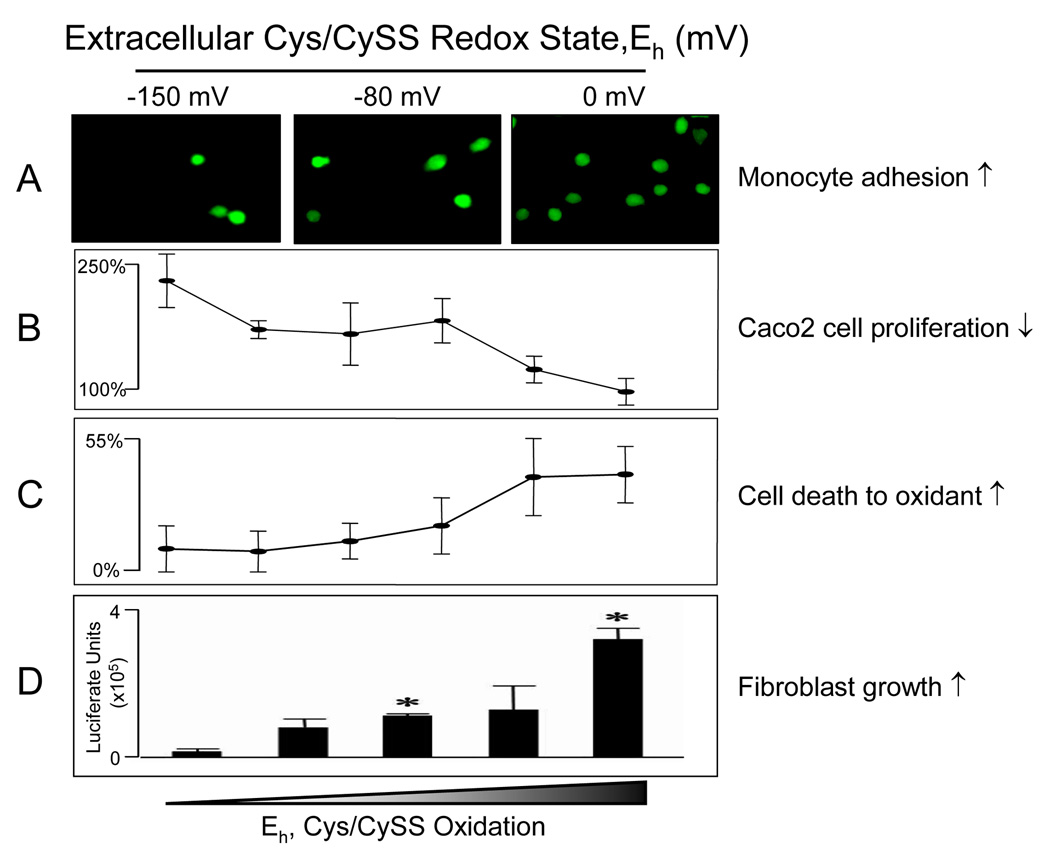
Diverse functions of eukaryotic cells are optimized by organization of compatible chemistries into distinct compartments defined by the structures of lipid-containing membranes, multiprotein complexes and oligomeric structures of saccharides and nucleic acids. This structural and chemical organization is coordinated, in part, through cysteine residues of proteins which undergo reversible oxidation-reduction and serve as chemical/structural transducing elements. The central thiol/disulfide redox couples, thioredoxin-1, thioredoxin-2, GSH/GSSG and cysteine/cystine (Cys/CySS), are not in equilibrium with each other and are maintained at distinct, non-equilibrium potentials in mitochondria, nuclei, the secretory pathway and the extracellular space. Mitochondria contain the most reducing compartment, have the highest rates of electron transfer and are highly sensitive to oxidation. Nuclei also have more reduced redox potentials but are relatively resistant to oxidation. The secretory pathway contains oxidative systems which introduce disulfides into proteins for export. The cytoplasm contains few metabolic oxidases and this maintains an environment for redox signaling dependent upon NADPH oxidases and NO synthases. Extracellular compartments are maintained at stable oxidizing potentials. Controlled changes in cytoplasmic GSH/GSSG redox potential are associated with functional state, varying with proliferation, differentiation and apoptosis. Variation in extracellular Cys/CySS redox potential is also associated with proliferation, cell adhesion and apoptosis. Thus, cellular redox biology is inseparable from redox compartmentalization. Further elucidation of the redox control networks within compartments will improve the mechanistic understanding of cell functions and their disruption in disease.
Figures

Similar articles
-
Oxidation of plasma cysteine/cystine redox state in endotoxin-induced lung injury.Am J Respir Cell Mol Biol. 2009 Jan;40(1):90-8. doi: 10.1165/rcmb.2007-0447OC. Epub 2008 Jul 29. Am J Respir Cell Mol Biol. 2009. PMID: 18664641 Free PMC article.
-
Intracellular proatherogenic events and cell adhesion modulated by extracellular thiol/disulfide redox state.Circulation. 2005 Jun 7;111(22):2973-80. doi: 10.1161/CIRCULATIONAHA.104.515155. Epub 2005 May 31. Circulation. 2005. PMID: 15927968
-
Age-dependent oxidation of extracellular cysteine/cystine redox state (Eh(Cys/CySS)) in mouse lung fibroblasts is mediated by a decline in Slc7a11 expression.Free Radic Biol Med. 2018 Apr;118:13-22. doi: 10.1016/j.freeradbiomed.2018.02.026. Epub 2018 Feb 16. Free Radic Biol Med. 2018. PMID: 29458149 Free PMC article.
-
Intestinal redox biology and oxidative stress.Semin Cell Dev Biol. 2012 Sep;23(7):729-37. doi: 10.1016/j.semcdb.2012.03.014. Epub 2012 Mar 30. Semin Cell Dev Biol. 2012. PMID: 22484611 Free PMC article. Review.
-
Redox compartmentalization and cellular stress.Diabetes Obes Metab. 2010 Oct;12 Suppl 2(Suppl 2):116-25. doi: 10.1111/j.1463-1326.2010.01266.x. Diabetes Obes Metab. 2010. PMID: 21029308 Free PMC article. Review.
Cited by
-
A redox switch regulates the assembly and anti-CRISPR activity of AcrIIC1.Nat Commun. 2022 Nov 18;13(1):7071. doi: 10.1038/s41467-022-34551-8. Nat Commun. 2022. PMID: 36400778 Free PMC article.
-
The intrinsically disordered membrane protein selenoprotein S is a reductase in vitro.Biochemistry. 2013 May 7;52(18):3051-61. doi: 10.1021/bi4001358. Epub 2013 Apr 24. Biochemistry. 2013. PMID: 23566202 Free PMC article.
-
Selenoprotein N in skeletal muscle: from diseases to function.J Mol Med (Berl). 2012 Oct;90(10):1095-107. doi: 10.1007/s00109-012-0896-x. Epub 2012 Apr 14. J Mol Med (Berl). 2012. PMID: 22527882 Review.
-
Bioactive VEGF-C from E. coli.Sci Rep. 2022 Oct 28;12(1):18157. doi: 10.1038/s41598-022-22960-0. Sci Rep. 2022. PMID: 36307539 Free PMC article.
-
Low-dose cadmium disrupts mitochondrial citric acid cycle and lipid metabolism in mouse lung.Free Radic Biol Med. 2019 Feb 1;131:209-217. doi: 10.1016/j.freeradbiomed.2018.12.005. Epub 2018 Dec 6. Free Radic Biol Med. 2019. PMID: 30529385 Free PMC article.
References
-
- Halvey PJ, Hansen JM, Lash LH, Jones DP. Compartmentation of Redox Signaling and Control: Discrimination of Oxidative Stress in Mitochondria, Cytoplasm. Nuclei and Endoplasmic Reticulum. 2007 In Press.
-
- Hansen JM, Go YM, Jones DP. Nuclear and mitochondrial compartmentation of oxidative stress and redox signaling. Annual review of pharmacology and toxicology. 2006;46:215–234. - PubMed
-
- Damdimopoulos AE, Miranda-Vizuete A, Pelto-Huikko M, Gustafsson JA, Spyrou G. Human mitochondrial thioredoxin. Involvement in mitochondrial membrane potential and cell death. The Journal of biological chemistry. 2002;277:33249–33257. - PubMed
-
- Watson WH, Pohl J, Montfort WR, Stuchlik O, Reed MS, Powis G, Jones DP. Redox potential of human thioredoxin 1 and identification of a second dithiol/disulfide motif. The Journal of biological chemistry. 2003;278:33408–33415. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
