

Bone morphogenetic protein signaling regulates the size of hair follicles and modulates the expression of cell cycle-associated genes
- PMID: 17114283
- PMCID: PMC1838724
- DOI: 10.1073/pnas.0608899103
Bone morphogenetic protein signaling regulates the size of hair follicles and modulates the expression of cell cycle-associated genes
Abstract
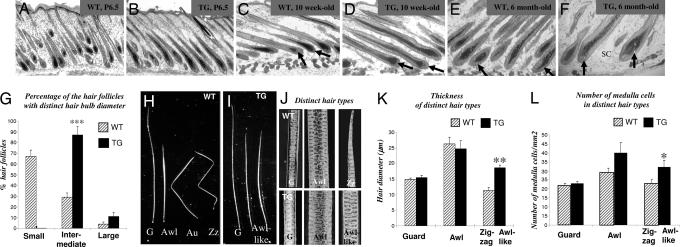
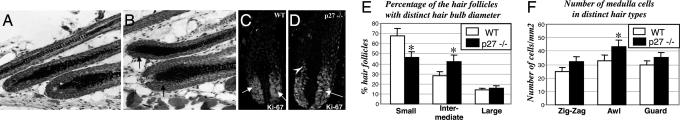
Bone morphogenetic protein (BMP) signaling is involved in the regulation of a large variety of developmental programs, including those controlling organ sizes. Here, we show that transgenic (TG) mice overexpressing the BMP antagonist noggin (promoter, K5) are characterized by a marked increase in size of anagen hair follicles (HFs) and by the replacement of zig-zag and auchen hairs by awl-like hairs, compared with the age-matched WT controls. Markedly enlarged anagen HFs of TG mice show increased proliferation in the matrix and an increased number of hair cortex and medulla cells compared with WT HFs. Microarray and real-time PCR analyses of the laser-captured hair matrix cells show a strong decrease in expression of Cdk inhibitor p27(Kip1) and increased expression of selected cyclins in TG vs. WT mice. Similar to TG mice, p27(Kip1) knockout mice also show an increased size of anagen HFs associated with increased cell proliferation in the hair bulb. Primary epidermal keratinocytes (KC) from TG mice exhibit significantly increased proliferation and decreased p27(Kip1) expression, compared with WT KC. Alternatively, activation of BMP signaling in HaCaT KC induces growth arrest, stimulates p27(Kip1) expression, and positively regulates p27(Kip1) promoter activity, thus further supporting a role of p27(Kip1) in mediating the effects of BMP signaling on HF size. These data suggest that BMP signaling plays an important role in regulating cell proliferation and controls the size of anagen HFs by modulating the expression of cell-cycle-associated genes in hair matrix KC.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


Similar articles
-
Characterisation of cell cycle arrest and terminal differentiation in a maximally proliferative human epithelial tissue: Lessons from the human hair follicle matrix.Eur J Cell Biol. 2017 Sep;96(6):632-641. doi: 10.1016/j.ejcb.2017.03.011. Epub 2017 Apr 2. Eur J Cell Biol. 2017. PMID: 28413121
-
Bone morphogenetic protein signaling regulates postnatal hair follicle differentiation and cycling.Am J Pathol. 2004 Sep;165(3):729-40. doi: 10.1016/S0002-9440(10)63336-6. Am J Pathol. 2004. PMID: 15331398 Free PMC article.
-
Bone morphogenetic protein signaling inhibits hair follicle anagen induction by restricting epithelial stem/progenitor cell activation and expansion.Stem Cells. 2006 Dec;24(12):2826-39. doi: 10.1634/stemcells.2005-0544. Epub 2006 Sep 7. Stem Cells. 2006. PMID: 16960130
-
Bone morphogenetic proteins and their antagonists in skin and hair follicle biology.J Invest Dermatol. 2003 Jan;120(1):36-47. doi: 10.1046/j.1523-1747.2003.12002.x. J Invest Dermatol. 2003. PMID: 12535196 Review.
-
Monstrous attempts at adnexogenesis: regulating hair follicle progenitors through Sonic hedgehog signaling.Curr Opin Genet Dev. 2001 Oct;11(5):541-6. doi: 10.1016/s0959-437x(00)00230-6. Curr Opin Genet Dev. 2001. PMID: 11532396 Review.
Cited by
-
p63 and Brg1 control developmentally regulated higher-order chromatin remodelling at the epidermal differentiation complex locus in epidermal progenitor cells.Development. 2014 Jan;141(1):101-11. doi: 10.1242/dev.103200. Development. 2014. PMID: 24346698 Free PMC article.
-
Bone morphogenetic protein signaling suppresses wound-induced skin repair by inhibiting keratinocyte proliferation and migration.J Invest Dermatol. 2014 Mar;134(3):827-837. doi: 10.1038/jid.2013.419. Epub 2013 Oct 14. J Invest Dermatol. 2014. PMID: 24126843 Free PMC article.
-
Whole-transcriptome gene expression profiling in an epidermolysis bullosa simplex Dowling-Meara model keratinocyte cell line uncovered novel, potential therapeutic targets and affected pathways.BMC Res Notes. 2015 Dec 15;8:785. doi: 10.1186/s13104-015-1783-7. BMC Res Notes. 2015. PMID: 26666517 Free PMC article.
-
Dermal papilla cell number specifies hair size, shape and cycling and its reduction causes follicular decline.Development. 2013 Apr;140(8):1676-83. doi: 10.1242/dev.090662. Epub 2013 Mar 13. Development. 2013. PMID: 23487317 Free PMC article.
-
Constitutively active Akt induces ectodermal defects and impaired bone morphogenetic protein signaling.Mol Biol Cell. 2008 Jan;19(1):137-49. doi: 10.1091/mbc.e07-08-0764. Epub 2007 Oct 24. Mol Biol Cell. 2008. PMID: 17959825 Free PMC article.
References
-
- Fuchs E, Raghavan S. Nat Rev Genet. 2002;3:199–209. - PubMed
-
- Fuchs E, Merrill BJ, Jamora C, DasGupta R. Dev Cell. 2001;1:13–25. - PubMed
-
- Millar SE. J Invest Dermatol. 2002;118:216–225. - PubMed
-
- Galbraith DB. Nature. 1969;222:288–290. - PubMed
-
- Sundberg JP, Hogan ME. Handbook of Mouse Mutations with Skin and Hair Abnormalities. Boca Raton, FL: CRC; 1994. pp. 57–68.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
