

Schizosaccharomyces pombe AGC family kinase Gad8p forms a conserved signaling module with TOR and PDK1-like kinases
- PMID: 12805221
- PMCID: PMC162150
- DOI: 10.1093/emboj/cdg298
Schizosaccharomyces pombe AGC family kinase Gad8p forms a conserved signaling module with TOR and PDK1-like kinases
Abstract
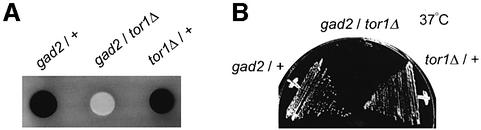
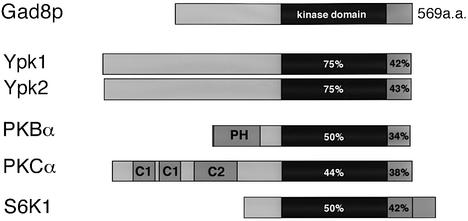
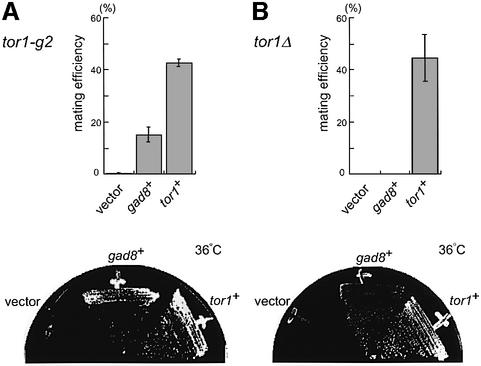
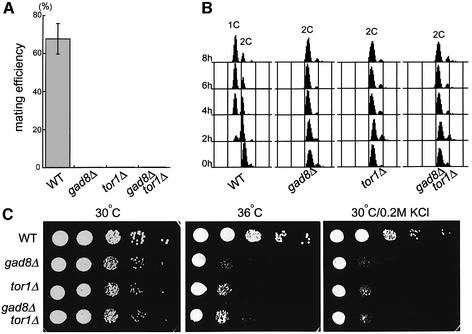
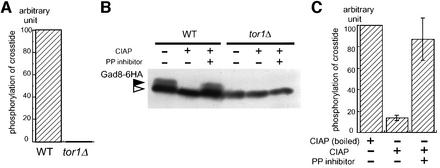
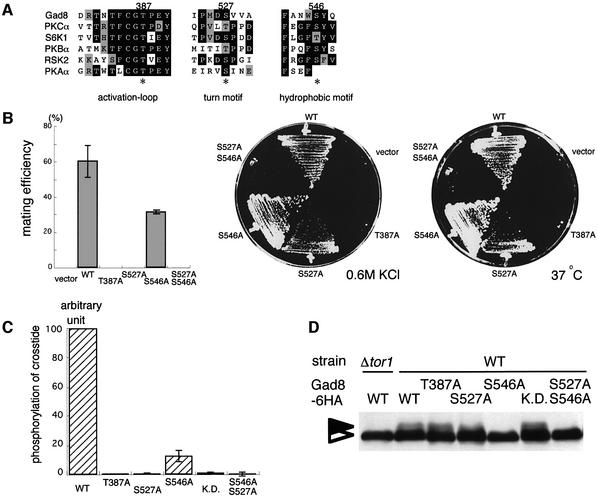
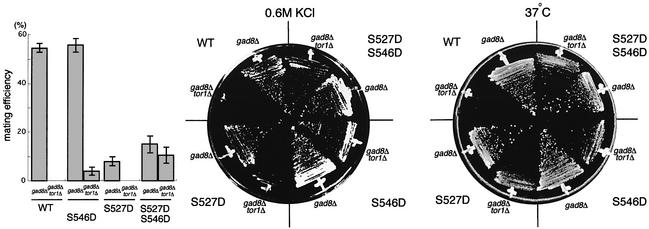
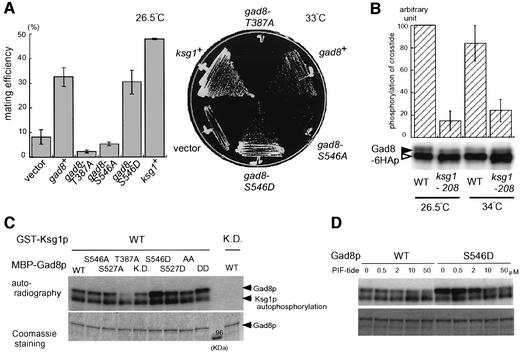
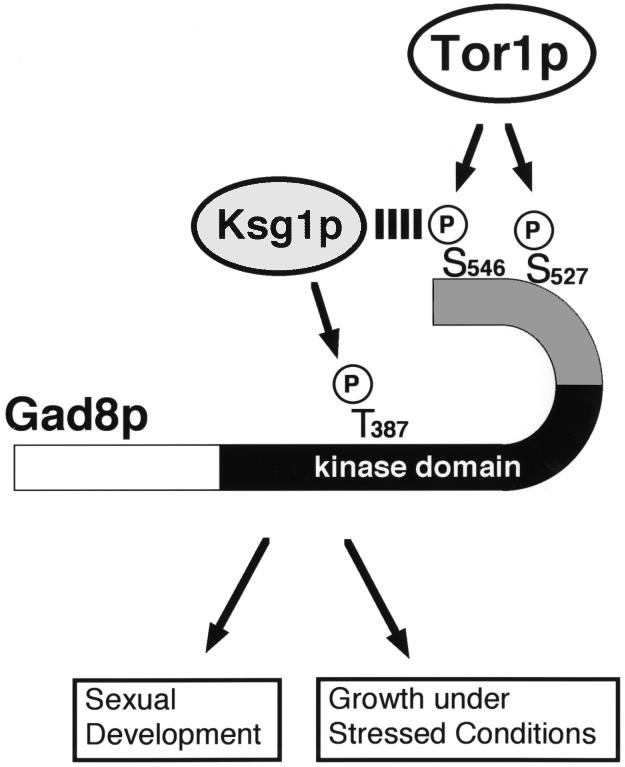
The TOR protein is a phosphoinositide kinase-related kinase widely conserved among eukaryotes. Fission yeast tor1 encodes an ortholog of TOR, which is required for sexual development and growth under stressed conditions. We isolated gad8, which encodes a Ser/Thr kinase of the AGC family, as a high-copy suppressor of the sterility of a tor1 mutant. Disruption of gad8 caused phenotypes similar to those of tor1 disruption. Gad8p was less phosphorylated and its kinase activity was undetectable in tor1Delta cells. Three amino acid residues corresponding to conserved phosphorylation sites in the AGC family kinases, namely Thr387 in the activation loop, Ser527 in the turn motif and Ser546 in the hydrophobic motif, were important for the kinase activity of Gad8p. Tor1p was responsible for the phosphorylation of Ser527 and Ser546, whereas Ksg1p, a PDK1-like kinase, appeared to phosphorylate Thr387 directly. Altogether, Tor1p, Ksg1p and Gad8p appear to constitute a signaling module for sexual development and growth under stressed conditions in fission yeast, which resembles the mTOR-PDK1-S6K1 system in mammals and may represent a basic signaling module ubiquitous in eukaryotes.
Figures
Similar articles
-
TOR Complex 2- independent mutations in the regulatory PIF pocket of Gad8AKT1/SGK1 define separate branches of the stress response mechanisms in fission yeast.PLoS Genet. 2020 Nov 2;16(11):e1009196. doi: 10.1371/journal.pgen.1009196. eCollection 2020 Nov. PLoS Genet. 2020. PMID: 33137119 Free PMC article.
-
Phosphoinositide-dependent kinase-1 orthologues from five eukaryotes are activated by the hydrophobic motif in AGC kinases.Biochem Biophys Res Commun. 2004 Sep 3;321(4):823-7. doi: 10.1016/j.bbrc.2004.07.031. Biochem Biophys Res Commun. 2004. PMID: 15358101
-
Psk1, an AGC kinase family member in fission yeast, is directly phosphorylated and controlled by TORC1 and functions as S6 kinase.J Cell Sci. 2012 Dec 1;125(Pt 23):5840-9. doi: 10.1242/jcs.111146. Epub 2012 Sep 12. J Cell Sci. 2012. PMID: 22976295 Free PMC article.
-
The spindle pole body plays a key role in controlling mitotic commitment in the fission yeast Schizosaccharomyces pombe.Biochem Soc Trans. 2008 Oct;36(Pt 5):1097-101. doi: 10.1042/BST0361097. Biochem Soc Trans. 2008. PMID: 18793196 Review.
-
PDK2: the missing piece in the receptor tyrosine kinase signaling pathway puzzle.Am J Physiol Endocrinol Metab. 2005 Aug;289(2):E187-96. doi: 10.1152/ajpendo.00011.2005. Am J Physiol Endocrinol Metab. 2005. PMID: 16014356 Review.
Cited by
-
Fission Yeast SCYL1/2 Homologue Ppk32: A Novel Regulator of TOR Signalling That Governs Survival during Brefeldin A Induced Stress to Protein Trafficking.PLoS Genet. 2016 May 18;12(5):e1006041. doi: 10.1371/journal.pgen.1006041. eCollection 2016 May. PLoS Genet. 2016. PMID: 27191590 Free PMC article.
-
TORC2 is required to maintain genome stability during S phase in fission yeast.J Biol Chem. 2013 Jul 5;288(27):19649-60. doi: 10.1074/jbc.M113.464974. Epub 2013 May 23. J Biol Chem. 2013. PMID: 23703609 Free PMC article.
-
TOR and MAP kinase pathways synergistically regulate autophagy in response to nutrient depletion in fission yeast.Autophagy. 2022 Feb;18(2):375-390. doi: 10.1080/15548627.2021.1935522. Epub 2021 Jun 23. Autophagy. 2022. PMID: 34157946 Free PMC article.
-
TORC2 and the AGC kinase Gad8 regulate phosphorylation of the ribosomal protein S6 in fission yeast.Biol Open. 2012 Sep 15;1(9):884-8. doi: 10.1242/bio.20122022. Epub 2012 Jul 17. Biol Open. 2012. PMID: 23213482 Free PMC article.
-
TORC1 Regulates Developmental Responses to Nitrogen Stress via Regulation of the GATA Transcription Factor Gaf1.mBio. 2015 Jul 7;6(4):e00959. doi: 10.1128/mBio.00959-15. mBio. 2015. PMID: 26152587 Free PMC article.
References
-
- Alessi D.R. (2001) Discovery of PDK1, one of the missing links in insulin signal transduction. Colworth Medal Lecture. Biochem. Soc. Trans, 29, 1–14. - PubMed
-
- Alessi D.R., Kozlowski,M.T., Weng,Q.P., Morrice,N. and Avruch,J. (1998) 3-Phosphoinositide-dependent protein kinase 1 (PDK1) phosphorylates and activates the p70 S6 kinase in vivo and in vitro. Curr. Biol., 8, 69–81. - PubMed
-
- Bahler J., Wu,J.Q., Longtine,M.S., Shah,N.G., McKenzie,A.,III, Steever,A.B., Wach,A., Philippsen,P. and Pringle,J.R. (1998) Heterologous modules for efficient and versatile PCR-based gene targeting in Schizosaccharomyces pombe. Yeast, 14, 943–951. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
