

Spectrum of Kaposi's sarcoma-associated herpesvirus, or human herpesvirus 8, diseases
- PMID: 12097251
- PMCID: PMC118087
- DOI: 10.1128/CMR.15.3.439-464.2002
Spectrum of Kaposi's sarcoma-associated herpesvirus, or human herpesvirus 8, diseases
Abstract
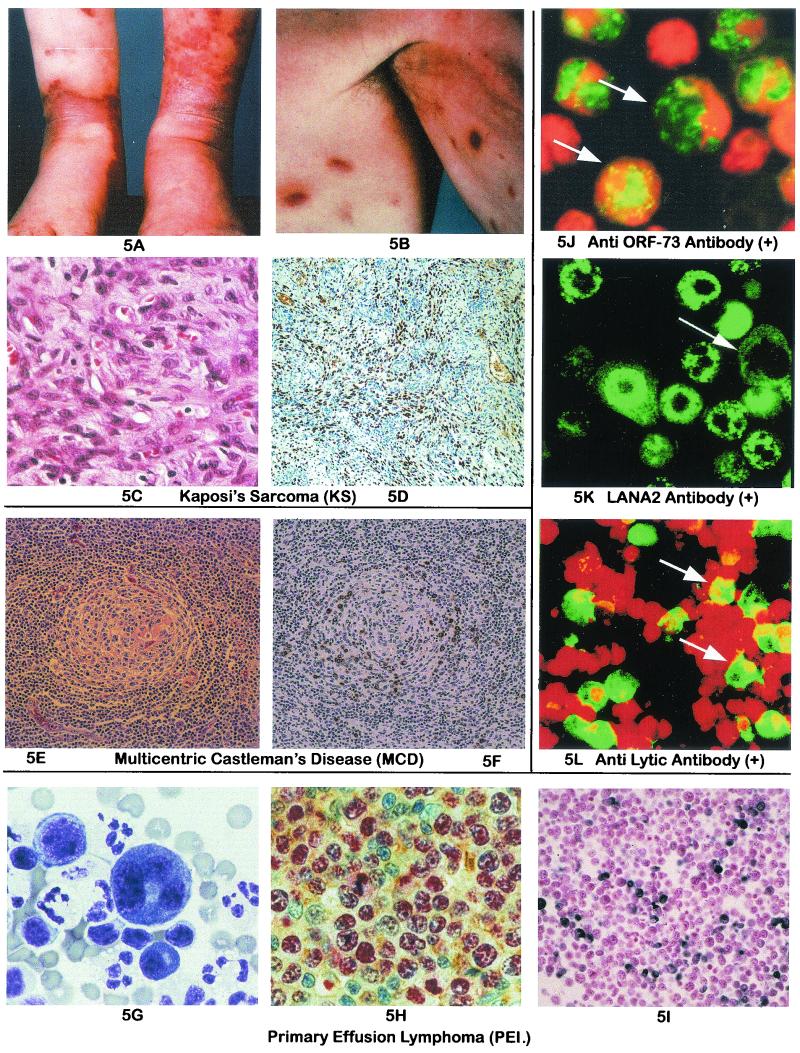
Human herpesvirus 8 (HHV-8), also known as Kaposi's sarcoma-associated herpesvirus (KSHV), discovered in 1994, is a human rhadinovirus (gamma-2 herpesvirus). Unlike other human herpesviruses (herpes simplex virus, Epstein-Barr virus, varicella-zoster virus, cytomegalovirus, HHV-6, and HHV-7), it is not widespread in the general population and has many unique proteins. HHV-8 is strongly associated with all subtypes of Kaposi's sarcoma (KS), multicentric Castleman's disease, and a rare form of B-cell lymphoma, primary effusion lymphoma. In addition, HHV-8 DNA sequences have been found in association with other diseases, but the role of the virus in these diseases is largely unconfirmed and remains controversial. The seroprevalence of HHV-8, based on detection of latent and lytic proteins, is 2 to 5% in healthy donors except in certain geographic areas where the virus is endemic, 80 to 95% in classic KS patients, and 40 to 50% in HIV-1 patients without KS. This virus can be transmitted both sexually and through body fluids (e.g., saliva and blood). HHV-8 is a transforming virus, as evidenced by its presence in human malignancies, by the in vitro transforming properties of several of its viral genes, and by its ability to transform some primary cells in culture. It is not, however, sufficient for transformation, and other cofactors such as immunosuppressive cytokines are involved in the development of HHV-8-associated malignancies. In this article, we review the biology, molecular virology, epidemiology, transmission, detection methods, pathogenesis, and antiviral therapy of this newly discovered human herpesvirus.
Figures
Similar articles
-
Kaposi's sarcoma-associated herpesvirus (human herpesvirus 8): epidemiology and pathogenesis.J Antimicrob Chemother. 2000 Apr;45 Suppl T3:15-27. doi: 10.1093/jac/45.suppl_4.15. J Antimicrob Chemother. 2000. PMID: 10855768 Review.
-
Human herpesvirus 8 (HHV-8) in the pathogenesis of Kaposi's sarcoma and other diseases.Leuk Lymphoma. 1998 Jan;28(3-4):255-64. doi: 10.3109/10428199809092681. Leuk Lymphoma. 1998. PMID: 9517497 Review.
-
[Human herpesvirus 8].Ann Med Interne (Paris). 1997;148(5):385-9. Ann Med Interne (Paris). 1997. PMID: 9538414 Review. French.
-
Kaposi's sarcoma and other manifestations of human herpesvirus 8.J Am Acad Dermatol. 2002 Nov;47(5):641-55; quiz 656-8. doi: 10.1067/mjd.2002.128383. J Am Acad Dermatol. 2002. PMID: 12399755 Review.
-
Characterization of the human herpesvirus 8 (Kaposi's sarcoma-associated herpesvirus) oncogene, kaposin (ORF K12).J Clin Virol. 2000 May;16(3):203-13. doi: 10.1016/s1386-6532(99)00081-5. J Clin Virol. 2000. PMID: 10738139
Cited by
-
Human herpesvirus 8: biology and role in the pathogenesis of Kaposi's sarcoma and other AIDS-related malignancies.Curr HIV/AIDS Rep. 2004 Apr;1(1):5-11. doi: 10.1007/s11904-004-0001-3. Curr HIV/AIDS Rep. 2004. PMID: 16091217 Review.
-
'RNA circles of influence' in Kaposi sarcoma.Ann Transl Med. 2019 Jul;7(Suppl 3):S109. doi: 10.21037/atm.2019.05.17. Ann Transl Med. 2019. PMID: 31576316 Free PMC article. No abstract available.
-
Human herpesvirus-8 infection in Tunisian adult acute leukemia patients.Afr Health Sci. 2023 Mar;23(1):504-510. doi: 10.4314/ahs.v23i1.52. Afr Health Sci. 2023. PMID: 37545962 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus genome programming during the early stages of primary infection of peripheral blood mononuclear cells.mBio. 2014 Dec 16;5(6):e02261-14. doi: 10.1128/mBio.02261-14. mBio. 2014. PMID: 25516617 Free PMC article.
-
Antimicrobial sulfonamides clear latent Kaposi sarcoma herpesvirus infection and impair MDM2-p53 complex formation.J Antibiot (Tokyo). 2017 Aug;70(9):962-966. doi: 10.1038/ja.2017.67. Epub 2017 Jun 14. J Antibiot (Tokyo). 2017. PMID: 28611469 Free PMC article.
References
-
- Ablashi, D. V., L. G. Chatlynne, H. C. Cooper, D. A. Thomas, M. Yadav, A. W. Norhanom, A. K. Chandana, V. Churdboonchart, S. A. R. Kulpradist, M. Patnaik, K. Liegmann, R. Masood, M. Reitz, F. Cleghorn, A. Manns, P. H. Levine, C. S. Rabkin, R. Biggar, F. Jensen, P. S. Gill, N. Jack, J. Edwards, J. E. Whitman, and C. Boshoff. 1999. Seroprevalence of human herpesvirus 8 (HHV-8) in countries of Southeast Asia compared to the United States, the Caribbean, and Africa. Br. J. Cancer 81:893-897. - PMC - PubMed
-
- Ablashi, D. V., L. Chatlynne, D. Thomas, D. Bourboulia, M. B. Rettig, R. A. Vescio, D. Viza, P. Gill, R. A. Kyle, J. R. Berenson, and J. E. Whitman, Jr. 2000. Lack of serologic association of human herpesvirus-8 (KSHV) in patients with monoclonal gammopathy of undetermined significance with and without progression to multiple myeloma. Blood 96:2304-2306. - PubMed
-
- Ablashi, D. V., L. Chatlynne, D. Thomas, C.-R. Lee, M. Roman, D. Bourboulia, S. Yoksan, C. Hall, R. Biggar, A. W. Norhanom, T. Yoshikawa, M. Margalith, and J. E. Whitman. 2000. Prevalence of HHV-8 (KSHV) latent and lytic IgG antibody in children: an international study. J. Hum. Virol. 3:261.
-
- Agbalika, F., X. Mariette, J.-P. Marolleau, J.-P. Fermand, and J.-C. Brouet. 1998. Detection of human herpesvirus-8 DNA in bone marrow biopsies from patients with multiple myeloma and Waldenstrom's macroglobulinemia. Blood 91:4393-4394. - PubMed
-
- Alexander, L., L. Denekamp, A. Knapp, M. R. Auerbach, B. Damania, and R. C. Desrosiers. 2000. The primary sequence of rhesus monkey rhadinovirus isolate 26-95: sequence similarities to Kaposi's sarcoma-associated herpesvirus and rhesus monkey rhadinovirus isolate 17577. J. Virol. 74:3388-3398. - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Medical
