

Epstein-Barr virus BALF1 is a BCL-2-like antagonist of the herpesvirus antiapoptotic BCL-2 proteins
- PMID: 11836425
- PMCID: PMC153809
- DOI: 10.1128/jvi.76.5.2469-2479.2002
Epstein-Barr virus BALF1 is a BCL-2-like antagonist of the herpesvirus antiapoptotic BCL-2 proteins
Abstract
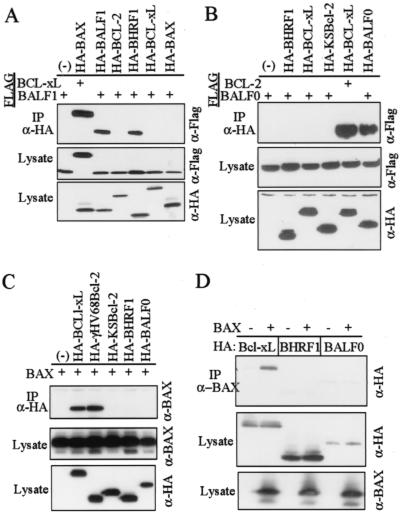
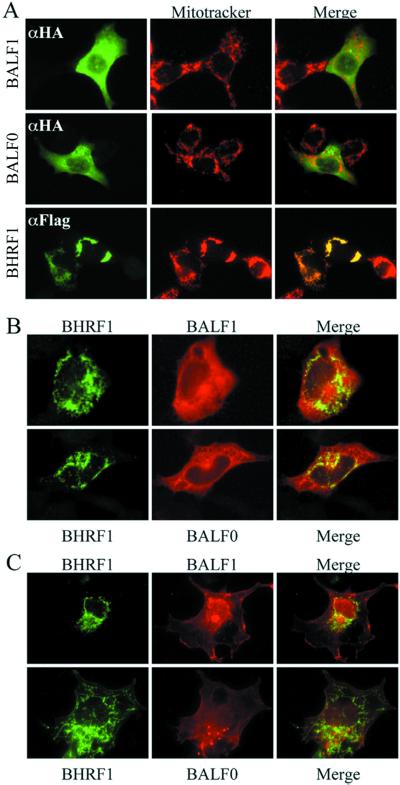
Cellular BCL-2 family proteins can inhibit or induce programmed cell death in part by counteracting the activity of other BCL-2 family members. All sequenced gammaherpesviruses encode a BCL-2 homologue that potently inhibits apoptosis and apparently escapes some of the regulatory mechanisms that govern the functions of their cellular counterparts. Examples of these protective proteins include BHRF1 of Epstein-Barr virus (EBV) and KSBcl-2 of Kaposi's sarcoma-associated herpesvirus, also known as human herpesvirus 8. The gamma-1 subgroup of these viruses, such as EBV, encodes a second BCL-2 homologue. We have now found that this second BCL-2 homologue encoded by EBV, BALF1, inhibits the antiapoptotic activity of EBV BHRF1 and of KSBcl-2 in several transfected cell lines. However, BALF1 failed to inhibit the cellular BCL-2 family member, BCL-x(L). Thus, BALF1 acts as a negative regulator of the survival function of BHRF1, similar to the counterbalance observed between cellular BCL-2 family members. Unlike the cellular BCL-2 family antagonists, BALF1 lacked proapoptotic activity and could not be converted into a proapoptotic factor in a manner similar to cellular BCL-2 proteins by caspase cleavage or truncation of the N terminus. Coimmunoprecipitation experiments and immunofluorescence assays suggest that a minimal amount, if any, of the BHRF1 and BALF1 proteins colocalizes inside cells, suggesting that mechanisms other than direct interaction explain the suppressive function of BALF1.
Figures

Similar articles
-
Antiapoptotic herpesvirus Bcl-2 homologs escape caspase-mediated conversion to proapoptotic proteins.J Virol. 2000 Jun;74(11):5024-31. doi: 10.1128/jvi.74.11.5024-5031.2000. J Virol. 2000. PMID: 10799576 Free PMC article.
-
Epstein-Barr virus encodes a novel homolog of the bcl-2 oncogene that inhibits apoptosis and associates with Bax and Bak.J Virol. 1999 Jun;73(6):5181-5. doi: 10.1128/JVI.73.6.5181-5185.1999. J Virol. 1999. PMID: 10233985 Free PMC article.
-
BHRF1, a viral homologue of the Bcl-2 oncogene, is conserved at both the sequence and functional level in different Epstein-Barr virus isolates.J Gen Virol. 1997 Nov;78 ( Pt 11):2987-99. doi: 10.1099/0022-1317-78-11-2987. J Gen Virol. 1997. PMID: 9367386
-
Viral versus cellular BCL-2 proteins.Cell Death Differ. 2003 Jan;10 Suppl 1:S68-76. doi: 10.1038/sj.cdd.4401133. Cell Death Differ. 2003. PMID: 12655348 Review.
-
Functional similarity between adenovirus E1B 19-kDa protein and proteins encoded by Bcl-2 proto-oncogene and Epstein-Barr virus BHRF1 gene.Curr Top Microbiol Immunol. 1995;199 ( Pt 1):153-61. doi: 10.1007/978-3-642-79496-4_9. Curr Top Microbiol Immunol. 1995. PMID: 7555053 Review. No abstract available.
Cited by
-
Epstein-Barr virus lytic gene BNRF1 promotes B-cell lymphomagenesis via IFI27 upregulation.PLoS Pathog. 2024 Feb 1;20(2):e1011954. doi: 10.1371/journal.ppat.1011954. eCollection 2024 Feb. PLoS Pathog. 2024. PMID: 38300891 Free PMC article.
-
Viral control of mitochondrial apoptosis.PLoS Pathog. 2008 May 30;4(5):e1000018. doi: 10.1371/journal.ppat.1000018. PLoS Pathog. 2008. PMID: 18516228 Free PMC article. Review.
-
Wildebeest-Derived Malignant Catarrhal Fever: A Bovine Peripheral T Cell Lymphoma Caused by Cross-Species Transmission of Alcelaphine Gammaherpesvirus 1.Viruses. 2023 Feb 13;15(2):526. doi: 10.3390/v15020526. Viruses. 2023. PMID: 36851740 Free PMC article. Review.
-
Viruses as modulators of mitochondrial functions.Adv Virol. 2013;2013:738794. doi: 10.1155/2013/738794. Epub 2013 Oct 24. Adv Virol. 2013. PMID: 24260034 Free PMC article. Review.
-
Role of the Crosstalk between Autophagy and Apoptosis in Cancer.J Oncol. 2013;2013:102735. doi: 10.1155/2013/102735. Epub 2013 Jun 5. J Oncol. 2013. PMID: 23840208 Free PMC article.
References
-
- Adams, J. M., and S. Cory. 1998. The Bcl-2 protein family: arbiters of cell survival. Science 281:1322-1326. - PubMed
-
- Basanez, G., A. Nechushtan, O. Drozhinin, A. Chanturiya, E. Choe, S. Tutt, K. A. Wood, Y.-T. Hsu, J. Zimmerberg, and R. J. Youle. 1999. Bax, but not Bcl-xL, decreases the lifetime of planar phospholipid bilayer membranes at subnanomolar concentrations. Proc. Natl. Acad. Sci. USA 96:5492-5497. - PMC - PubMed
-
- Basanez, G., J. Zhang, B. N. Chau, G. I. Maksaev, V. Frolov, T. A. Brandt, J. Burch, J. M. Hardwick, and J. Zimmerberg. 2001. Pro-apoptotic cleavage products of Bcl-xL form cytochrome c-conducting pores in pure lipid bilayers. J. Biol. Chem. 276:31083-31091. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
