

E-cadherin regulates cell growth by modulating proliferation-dependent beta-catenin transcriptional activity
- PMID: 11564756
- PMCID: PMC2150811
- DOI: 10.1083/jcb.200104036
E-cadherin regulates cell growth by modulating proliferation-dependent beta-catenin transcriptional activity
Abstract
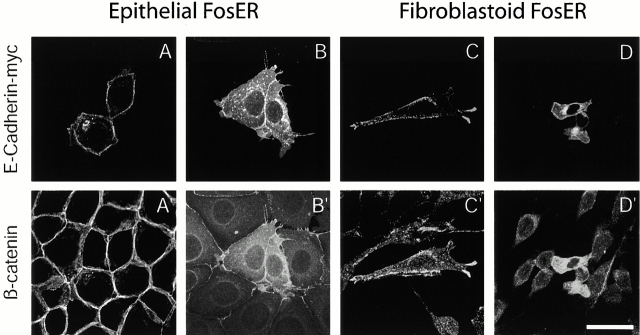
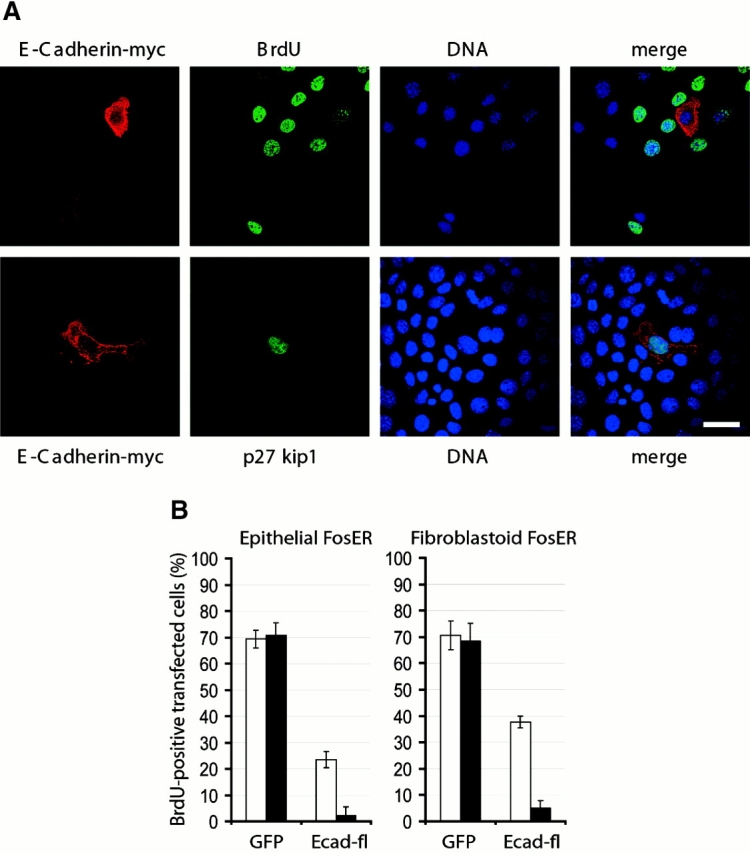
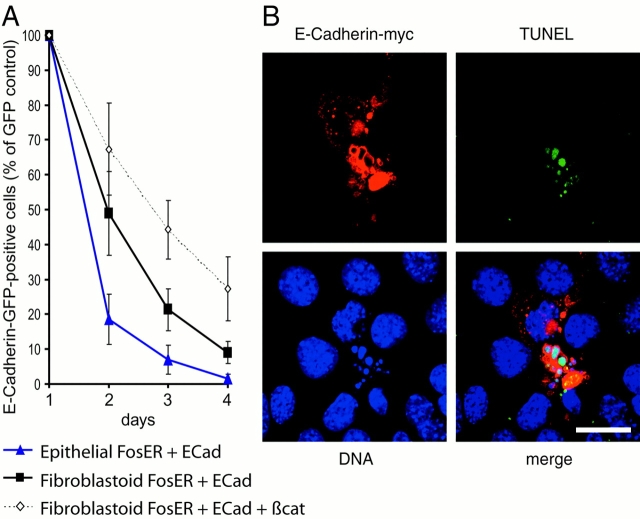
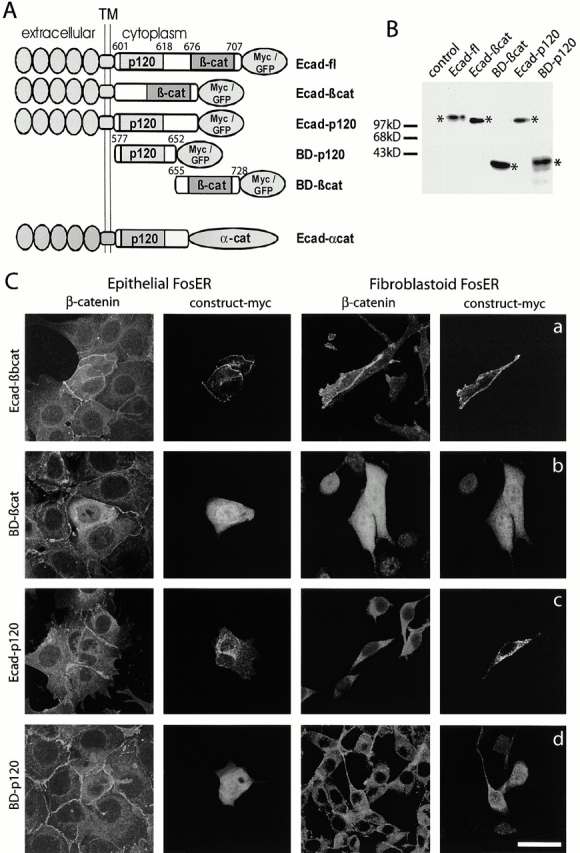
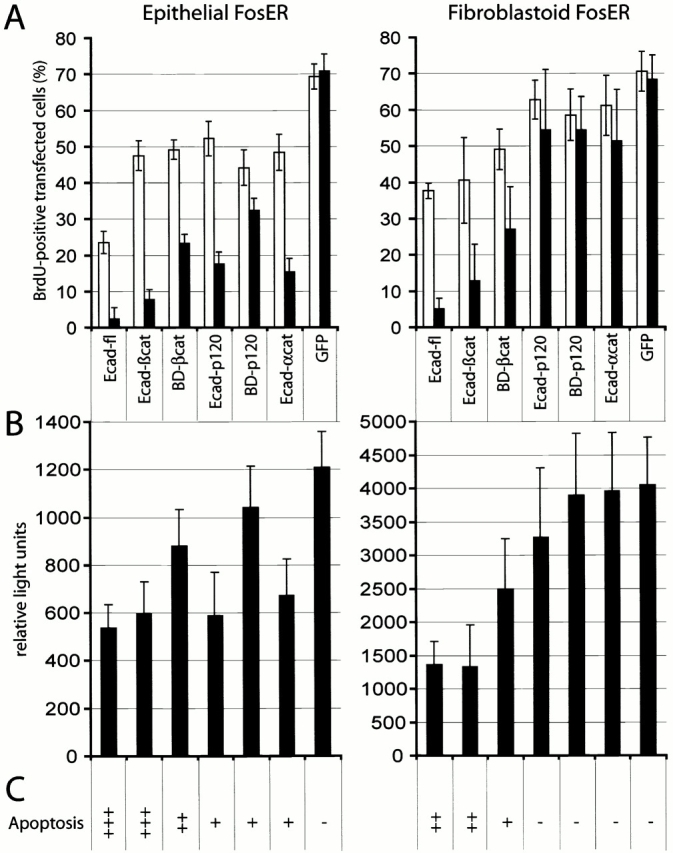
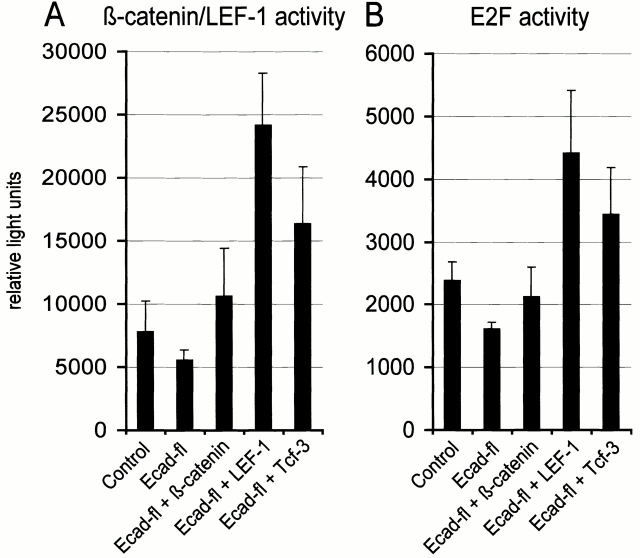
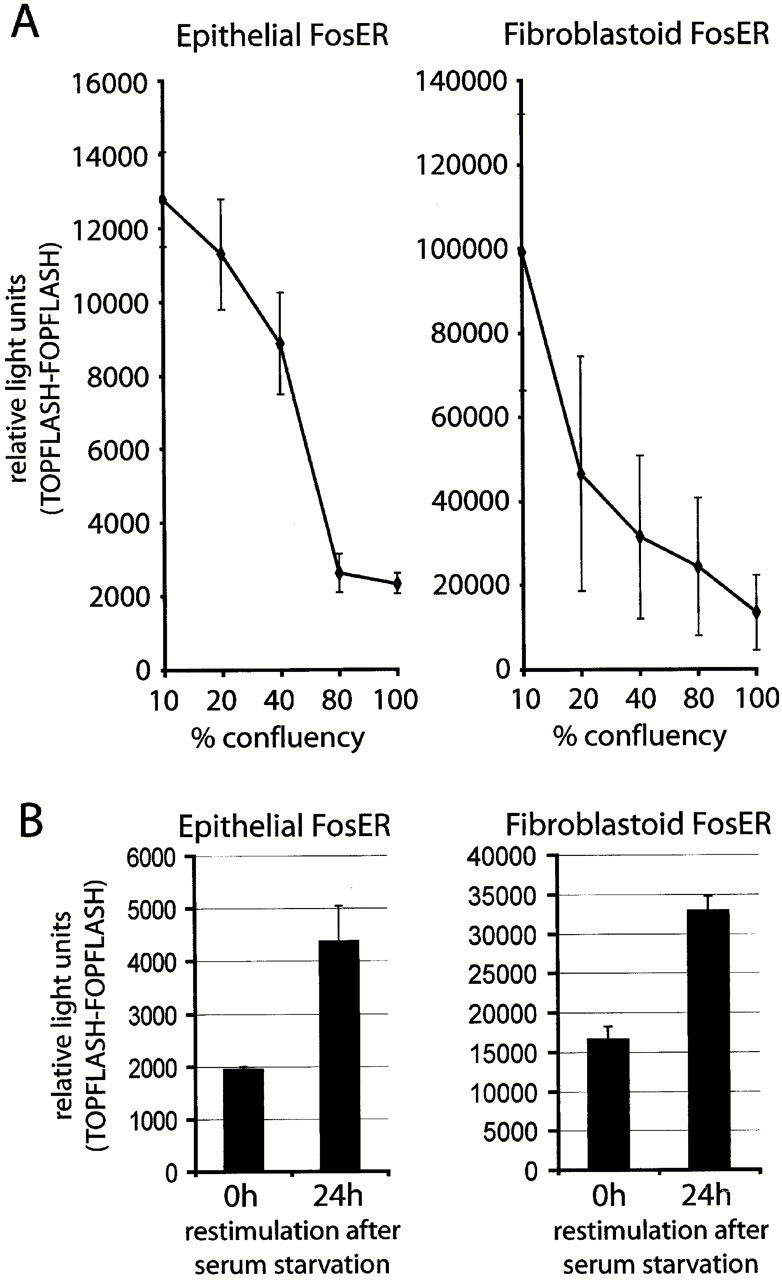
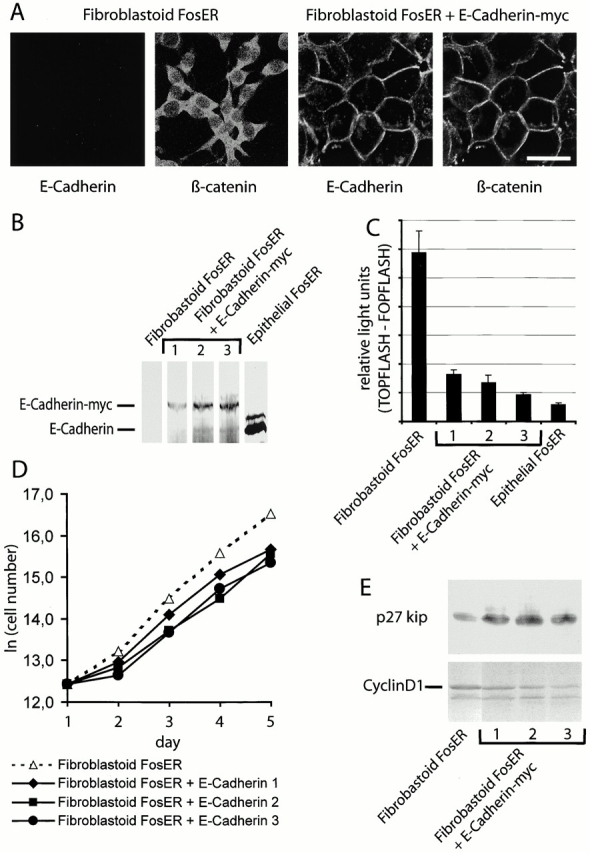
beta-Catenin is essential for E-cadherin-mediated cell adhesion in epithelial cells, but it also forms nuclear complexes with high mobility group transcription factors. Using a mouse mammary epithelial cell system, we have shown previously that conversion of epithelial cells to a fibroblastoid phenotype (epithelial-mesenchymal transition) involves downregulation of E-cadherin and upregulation of beta-catenin transcriptional activity. Here, we demonstrate that transient expression of exogenous E-cadherin in both epithelial and fibroblastoid cells arrested cell growth or caused apoptosis, depending on the cellular E-cadherin levels. By expressing E-cadherin subdomains, we show that the growth-suppressive effect of E-cadherin required the presence of its cytoplasmic beta-catenin interaction domain and/or correlated strictly with the ability to negatively interfere with beta-catenin transcriptional activity. Furthermore, coexpression of beta-catenin or lymphoid enhancer binding factor-1 or T cell factor 3 with E-cadherin rescued beta-catenin transcriptional activity and counteracted E-cadherin-mediated cell cycle arrest. Stable expression of E-cadherin in fibroblastoid cells decreased beta-catenin activity and reduced cell growth. Since proliferating cells had a higher beta-catenin activity than G1 phase-arrested or contact-inhibited cells, we conclude that beta-catenin transcriptional activity is essential for cell proliferation and can be controlled by E-cadherin in a cell adhesion-independent manner.
Figures
Similar articles
-
Epithelial mesenchymal transition by c-Fos estrogen receptor activation involves nuclear translocation of beta-catenin and upregulation of beta-catenin/lymphoid enhancer binding factor-1 transcriptional activity.J Cell Biol. 2000 Jan 10;148(1):173-88. doi: 10.1083/jcb.148.1.173. J Cell Biol. 2000. PMID: 10629227 Free PMC article.
-
E-cadherin suppresses cellular transformation by inhibiting beta-catenin signaling in an adhesion-independent manner.J Cell Biol. 2001 May 28;153(5):1049-60. doi: 10.1083/jcb.153.5.1049. J Cell Biol. 2001. PMID: 11381089 Free PMC article.
-
beta-Catenin and TGFbeta signalling cooperate to maintain a mesenchymal phenotype after FosER-induced epithelial to mesenchymal transition.Oncogene. 2004 Apr 8;23(15):2672-2680. doi: 10.1038/sj.onc.1207416. Oncogene. 2004. PMID: 14755243
-
The cadherin-catenin complex as a focal point of cell adhesion and signalling: new insights from three-dimensional structures.Bioessays. 2004 May;26(5):497-511. doi: 10.1002/bies.20033. Bioessays. 2004. PMID: 15112230 Review.
-
Defective E-cadherin/catenin complexes in human cancer.Virchows Arch. 2001 Dec;439(6):725-51. doi: 10.1007/s004280100516. Virchows Arch. 2001. PMID: 11787845 Review.
Cited by
-
Galectin-1 and Galectin-3 Mediate Protocadherin-24-Dependent Membrane Localization of β-catenin in Colon Cancer Cell Line HCT116.Curr Chem Genomics. 2012;6:18-26. doi: 10.2174/1875397301206010018. Epub 2012 Sep 20. Curr Chem Genomics. 2012. PMID: 23115611 Free PMC article.
-
hNaa10p contributes to tumorigenesis by facilitating DNMT1-mediated tumor suppressor gene silencing.J Clin Invest. 2010 Aug;120(8):2920-30. doi: 10.1172/JCI42275. Epub 2010 Jul 1. J Clin Invest. 2010. PMID: 20592467 Free PMC article.
-
Proliferation, but not apoptosis, is associated with distinct beta-catenin expression patterns in non-small-cell lung carcinomas: relationship with adenomatous polyposis coli and G(1)-to S-phase cell-cycle regulators.Am J Pathol. 2002 Nov;161(5):1619-34. doi: 10.1016/s0002-9440(10)64440-9. Am J Pathol. 2002. PMID: 12414510 Free PMC article.
-
Transcriptional modulation of TCR, Notch and Wnt signaling pathways in SEB-anergized CD4+ T cells.Genes Immun. 2005 Oct;6(7):596-608. doi: 10.1038/sj.gene.6364245. Genes Immun. 2005. PMID: 16034473 Free PMC article.
-
Distinct cholangiocarcinoma cell migration in 2D monolayer and 3D spheroid culture based on galectin-3 expression and localization.Front Oncol. 2023 Jan 12;12:999158. doi: 10.3389/fonc.2022.999158. eCollection 2022. Front Oncol. 2023. PMID: 36713574 Free PMC article.
References
-
- Anastasiadis, P.Z., and A.B. Reynolds. 2000. The p120 catenin family: complex roles in adhesion, signaling and cancer. J. Cell Sci. 113:1319–1334. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
