

Binding of human immunodeficiency virus type 1 gp120 to CXCR4 induces mitochondrial transmembrane depolarization and cytochrome c-mediated apoptosis independently of Fas signaling
- PMID: 11462036
- PMCID: PMC114999
- DOI: 10.1128/JVI.75.16.7637-7650.2001
Binding of human immunodeficiency virus type 1 gp120 to CXCR4 induces mitochondrial transmembrane depolarization and cytochrome c-mediated apoptosis independently of Fas signaling
Abstract
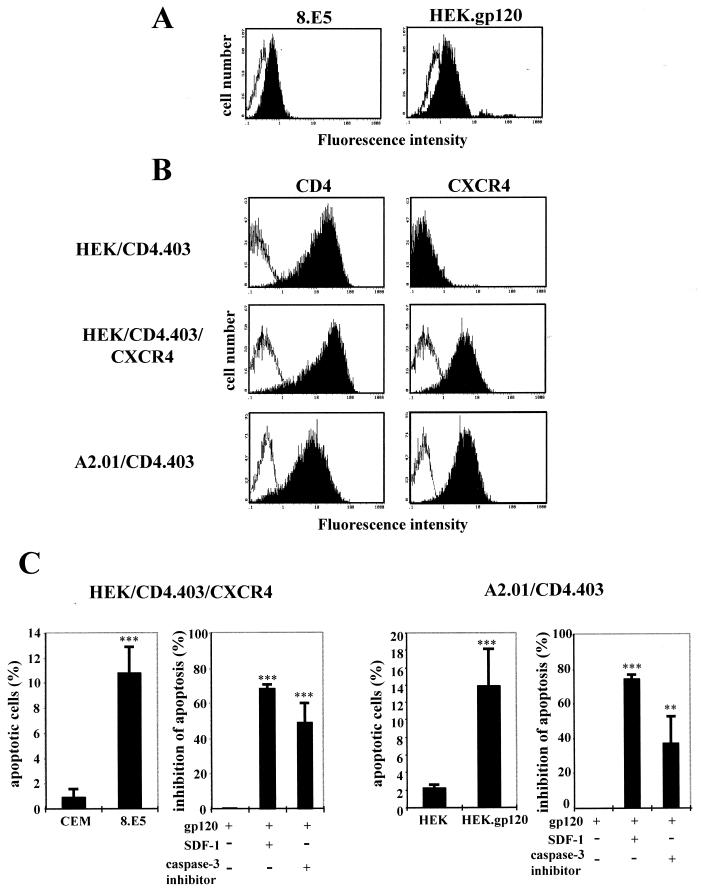
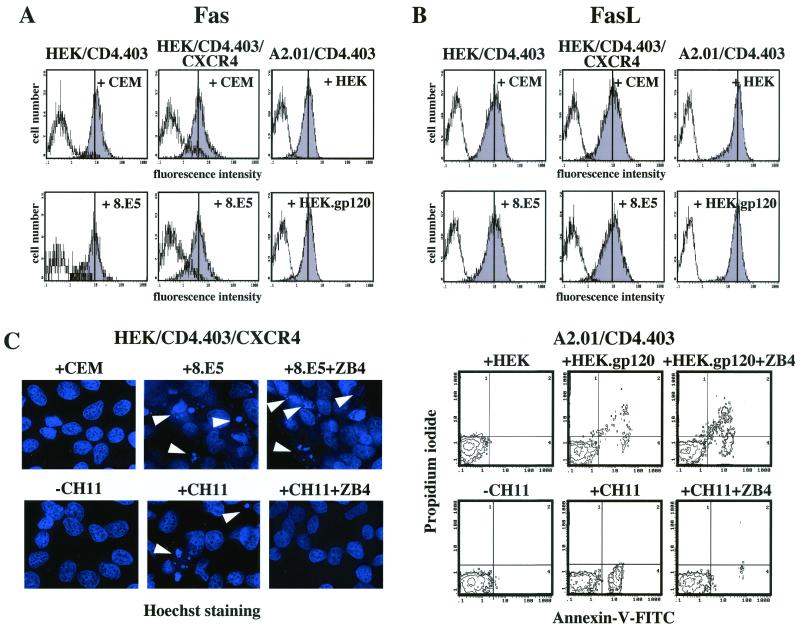
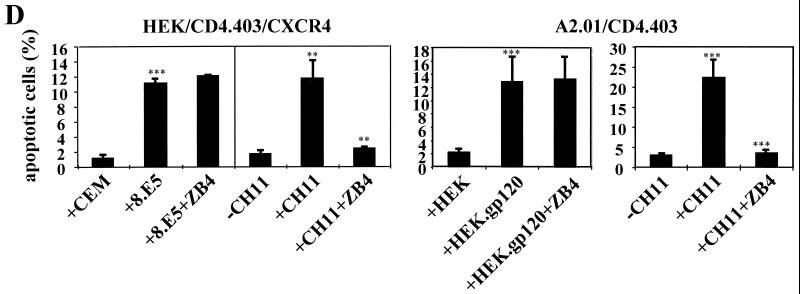
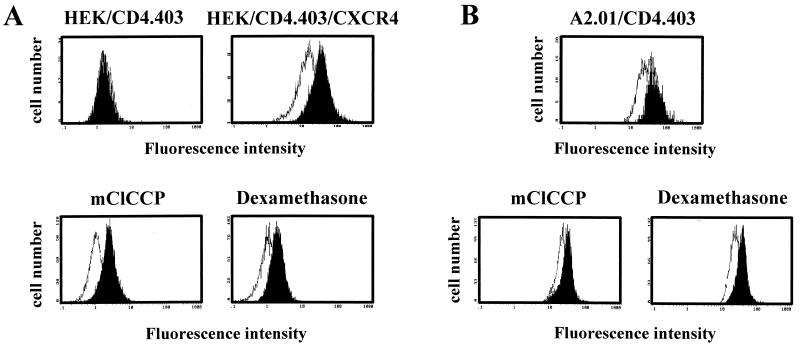
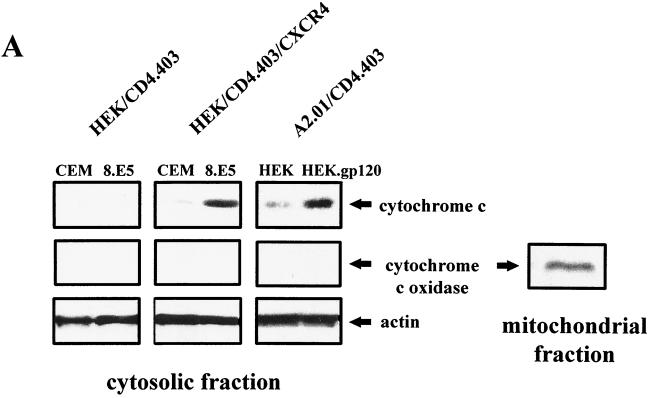
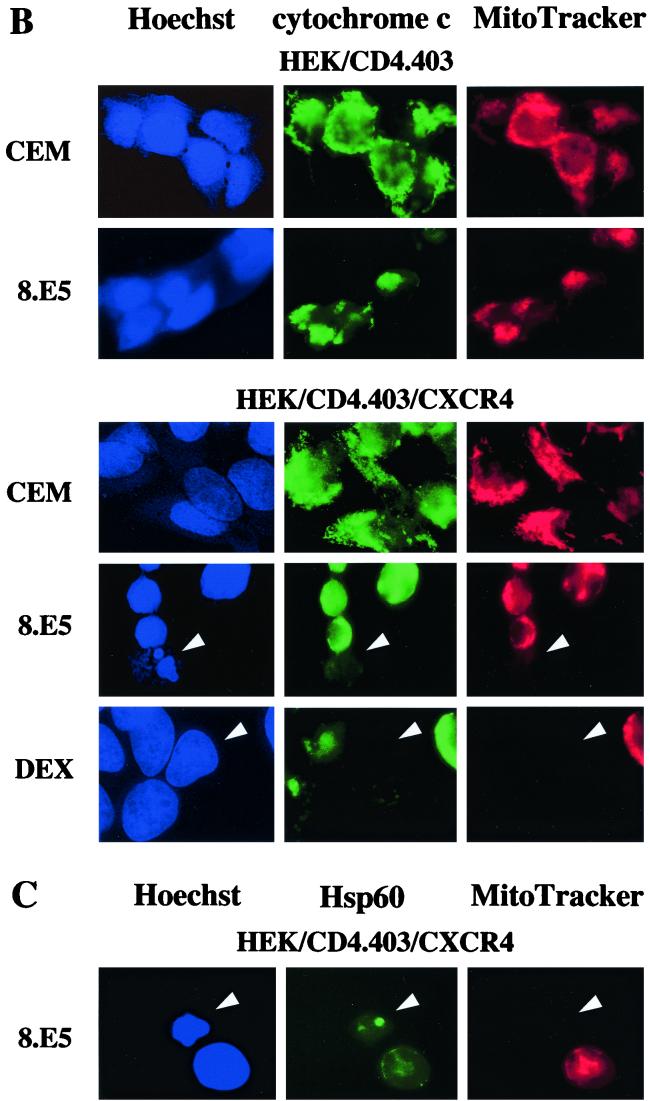
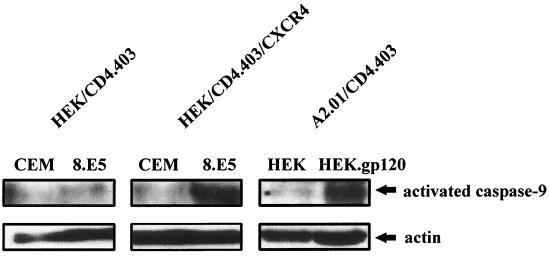
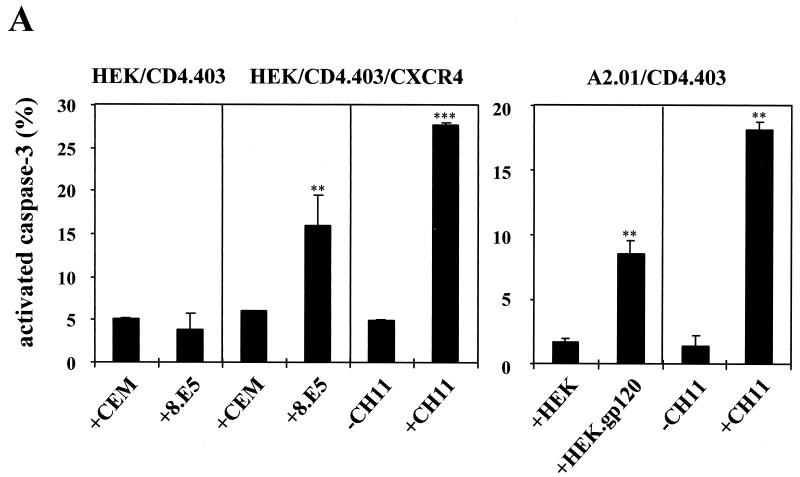
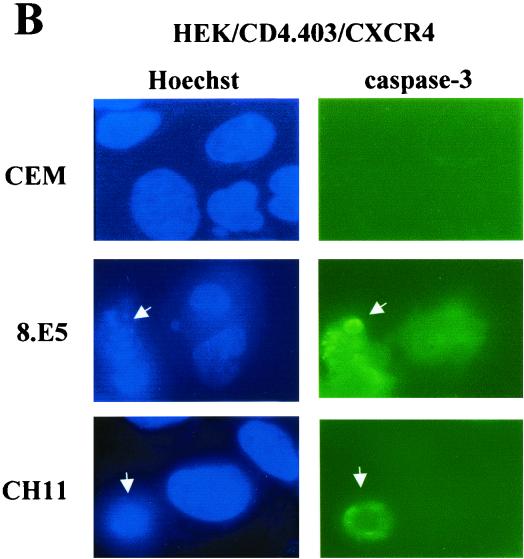
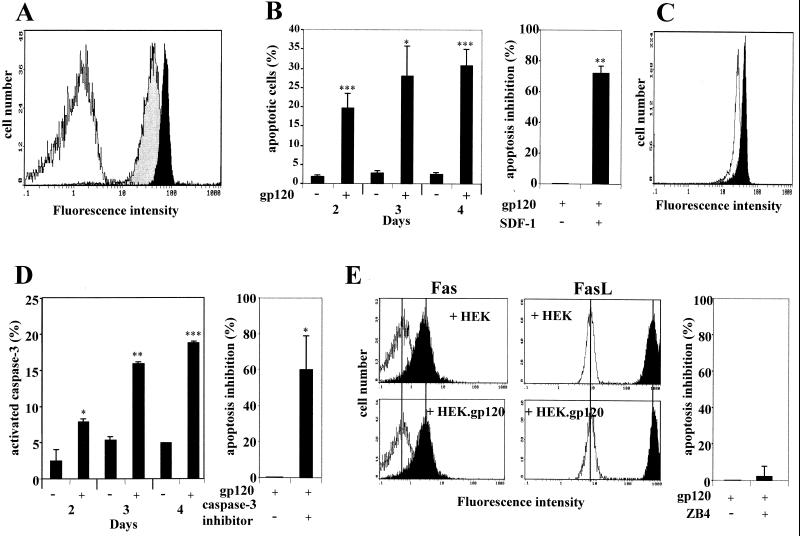
Apoptosis of CD4(+) T lymphocytes, induced by contact between human immunodeficiency virus type 1 (HIV-1) envelope glycoprotein (gp120) and its receptors, could contribute to the cell depletion observed in HIV-infected individuals. CXCR4 appears to play an important role in gp120-induced cell death, but the mechanisms involved in this apoptotic process remain poorly understood. To get insight into the signal transduction pathways connecting CXCR4 to apoptosis following gp120 binding, we used different cell lines expressing wild-type CXCR4 and a truncated form of CD4 that binds gp120 but lacks the ability to transduce signals. The present study demonstrates that (i) the interaction of cell-associated gp120 with CXCR4-expressing target cells triggers a rapid dissipation of the mitochondrial transmembrane potential resulting in the cytosolic release of cytochrome c from the mitochondria to cytosol, concurrent with activation of caspase-9 and -3; (ii) this apoptotic process is independent of Fas signaling; and (iii) cooperation with a CD4 signal is not required. In addition, following coculture with cells expressing gp120, a Fas-independent apoptosis involving mitochondria and caspase activation is also observed in primary umbilical cord blood CD4(+) T lymphocytes expressing high levels of CXCR4. Thus, this gp120-mediated apoptotic pathway may contribute to CD4(+) T-cell depletion in AIDS.
Figures
Similar articles
-
Caspase-dependent apoptosis of cells expressing the chemokine receptor CXCR4 is induced by cell membrane-associated human immunodeficiency virus type 1 envelope glycoprotein (gp120).Virology. 2000 Mar 15;268(2):329-44. doi: 10.1006/viro.1999.0151. Virology. 2000. PMID: 10704341
-
HIV envelope glycoprotein-induced cell killing by apoptosis is enhanced with increased expression of CD26 in CD4+ T cells.Virology. 1996 Sep 15;223(2):318-30. doi: 10.1006/viro.1996.0483. Virology. 1996. PMID: 8806567
-
Mechanisms of apoptosis induction by the HIV-1 envelope.Cell Death Differ. 2005 Aug;12 Suppl 1:916-23. doi: 10.1038/sj.cdd.4401584. Cell Death Differ. 2005. PMID: 15719026 Review.
-
Apoptosis of CD8+ T cells is mediated by macrophages through interaction of HIV gp120 with chemokine receptor CXCR4.Nature. 1998 Sep 10;395(6698):189-94. doi: 10.1038/26026. Nature. 1998. PMID: 9744279
-
Are blockers of gp120/CD4 interaction effective inhibitors of HIV-1 immunopathogenesis?AIDS Rev. 2006 Jan-Mar;8(1):3-8. AIDS Rev. 2006. PMID: 16736946 Review.
Cited by
-
Reactive Oxygen Species (ROS) are Critical for Morphine Exacerbation of HIV-1 gp120-Induced Pain.J Neuroimmune Pharmacol. 2021 Sep;16(3):581-591. doi: 10.1007/s11481-020-09951-6. Epub 2020 Aug 22. J Neuroimmune Pharmacol. 2021. PMID: 32827051 Free PMC article.
-
Sequential involvement of Cdk1, mTOR and p53 in apoptosis induced by the HIV-1 envelope.EMBO J. 2002 Aug 1;21(15):4070-80. doi: 10.1093/emboj/cdf391. EMBO J. 2002. PMID: 12145207 Free PMC article.
-
HIV gp120 induces, NF-kappaB dependent, HIV replication that requires procaspase 8.PLoS One. 2009;4(3):e4875. doi: 10.1371/journal.pone.0004875. Epub 2009 Mar 16. PLoS One. 2009. PMID: 19287489 Free PMC article.
-
The interaction between mitochondria and oncoviruses.Biochim Biophys Acta Mol Basis Dis. 2018 Feb;1864(2):481-487. doi: 10.1016/j.bbadis.2017.09.023. Epub 2017 Sep 28. Biochim Biophys Acta Mol Basis Dis. 2018. PMID: 28962899 Free PMC article. Review.
-
HIV-1 Env induces pexophagy and an oxidative stress leading to uninfected CD4+ T cell death.Autophagy. 2021 Sep;17(9):2465-2474. doi: 10.1080/15548627.2020.1831814. Epub 2020 Oct 19. Autophagy. 2021. PMID: 33073673 Free PMC article.
References
-
- Adachi S, Gottlieb R A, Babior B M. Lack of release of cytochrome C from mitochondria into cytosol early in the course of Fas-mediated apoptosis of Jurkat cells. J Biol Chem. 1998;273:19892–19894. - PubMed
-
- Badley A D, Dockrell D, Paya C V. Apoptosis in AIDS. Adv Pharmacol. 1997;41:271–294. - PubMed
-
- Badley A D, Parato K, Cameron D W, Kravcik S, Phenix B N, Ashby D, Kumar A, Lynch D H, Tschopp J, Angel J B. Dynamic correlation of apoptosis and immune activation during treatment of HIV infection. Cell Death Differ. 1999;6:420–432. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
