

Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation
- PMID: 11015443
- PMCID: PMC2193311
- DOI: 10.1084/jem.192.7.1027
Engagement of the PD-1 immunoinhibitory receptor by a novel B7 family member leads to negative regulation of lymphocyte activation
Abstract
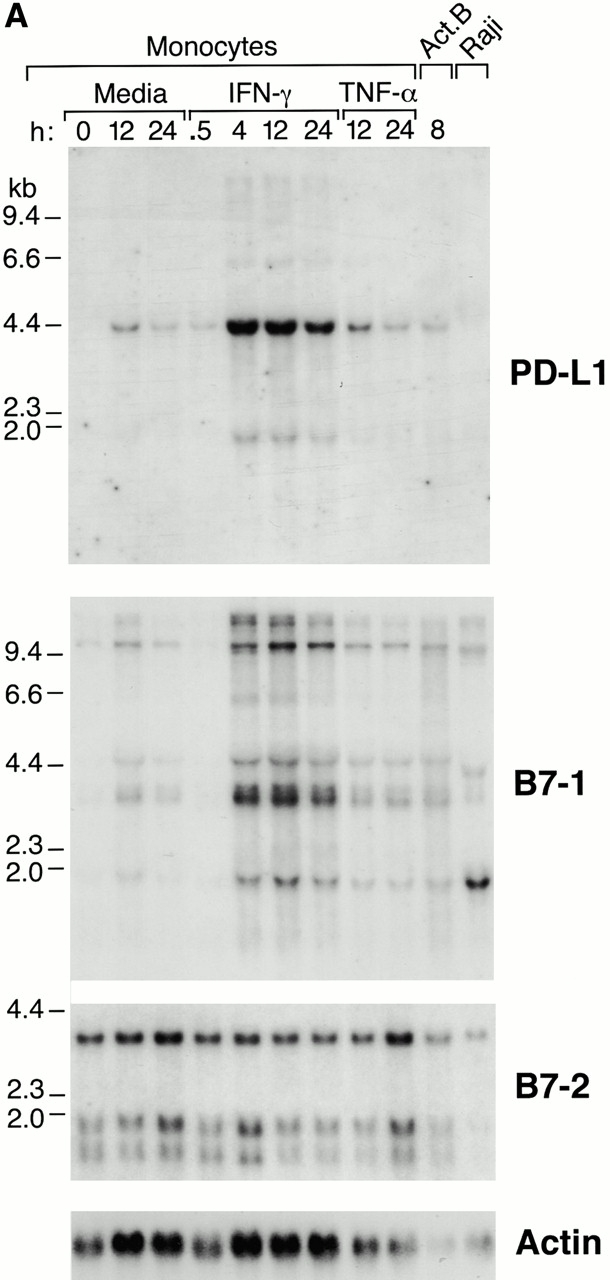
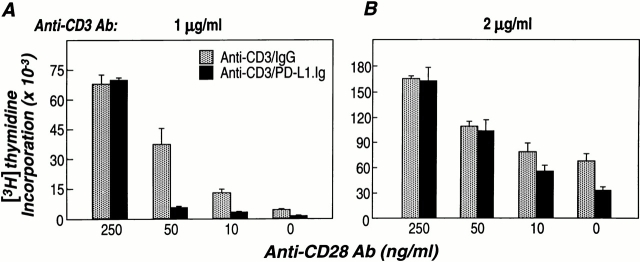
PD-1 is an immunoinhibitory receptor expressed by activated T cells, B cells, and myeloid cells. Mice deficient in PD-1 exhibit a breakdown of peripheral tolerance and demonstrate multiple autoimmune features. We report here that the ligand of PD-1 (PD-L1) is a member of the B7 gene family. Engagement of PD-1 by PD-L1 leads to the inhibition of T cell receptor-mediated lymphocyte proliferation and cytokine secretion. In addition, PD-1 signaling can inhibit at least suboptimal levels of CD28-mediated costimulation. PD-L1 is expressed by antigen-presenting cells, including human peripheral blood monocytes stimulated with interferon gamma, and activated human and murine dendritic cells. In addition, PD-L1 is expressed in nonlymphoid tissues such as heart and lung. The relative levels of inhibitory PD-L1 and costimulatory B7-1/B7-2 signals on antigen-presenting cells may determine the extent of T cell activation and consequently the threshold between tolerance and autoimmunity. PD-L1 expression on nonlymphoid tissues and its potential interaction with PD-1 may subsequently determine the extent of immune responses at sites of inflammation.
Figures









Similar articles
-
Analysis of the role of negative T cell costimulatory pathways in CD4 and CD8 T cell-mediated alloimmune responses in vivo.J Immunol. 2005 Jun 1;174(11):6648-56. doi: 10.4049/jimmunol.174.11.6648. J Immunol. 2005. PMID: 15905503
-
Expression of programmed death 1 ligands by murine T cells and APC.J Immunol. 2002 Nov 15;169(10):5538-45. doi: 10.4049/jimmunol.169.10.5538. J Immunol. 2002. PMID: 12421930
-
PD-1:PD-L inhibitory pathway affects both CD4(+) and CD8(+) T cells and is overcome by IL-2.Eur J Immunol. 2002 Mar;32(3):634-43. doi: 10.1002/1521-4141(200203)32:3<634::AID-IMMU634>3.0.CO;2-9. Eur J Immunol. 2002. PMID: 11857337
-
The expanding world of co-stimulation: the two-signal model revisited.Trends Immunol. 2001 Apr;22(4):217-23. doi: 10.1016/s1471-4906(01)01868-3. Trends Immunol. 2001. PMID: 11274928 Review.
-
Negative regulation of T-cell function by PD-1.Crit Rev Immunol. 2004;24(4):229-37. doi: 10.1615/critrevimmunol.v24.i4.10. Crit Rev Immunol. 2004. PMID: 15588223 Review.
Cited by
-
Anti-angiogenic agents - overcoming tumour endothelial cell anergy and improving immunotherapy outcomes.Nat Rev Clin Oncol. 2021 Aug;18(8):527-540. doi: 10.1038/s41571-021-00496-y. Epub 2021 Apr 8. Nat Rev Clin Oncol. 2021. PMID: 33833434 Review.
-
Lipid metabolic features of T cells in the Tumor Microenvironment.Lipids Health Dis. 2022 Oct 6;21(1):94. doi: 10.1186/s12944-022-01705-y. Lipids Health Dis. 2022. PMID: 36203151 Free PMC article. Review.
-
The efficacy and safety of antibodies targeting PD-1 for treatment in advanced esophageal cancer: A systematic review and meta-analysis.Transl Oncol. 2021 Jun;14(6):101083. doi: 10.1016/j.tranon.2021.101083. Epub 2021 Mar 27. Transl Oncol. 2021. PMID: 33784583 Free PMC article.
-
Novel immunotherapies in GU malignancies.Curr Oncol Rep. 2013 Jun;15(3):224-31. doi: 10.1007/s11912-013-0306-8. Curr Oncol Rep. 2013. PMID: 23519772 Review.
-
Long-term Benefit of PD-L1 Blockade in Lung Cancer Associated with JAK3 Activation.Cancer Immunol Res. 2015 Aug;3(8):855-63. doi: 10.1158/2326-6066.CIR-15-0024. Epub 2015 May 26. Cancer Immunol Res. 2015. PMID: 26014096 Free PMC article.
References
-
- Agata Y., Kawasaki A., Nishimura H., Ishida Y., Tsubata T., Yagita H., Honjo T. Expression of the PD-1 antigen on the surface of stimulated mouse T and B lymphocytes. Int. Immunol. 1996;8:765–772. - PubMed
-
- Vibhakar R., Juan G., Traganos F., Darzynkiewicz Z., Finger L.R. Activation-induced expression of human programmed death-1 gene in T-lymphocytes. Exp. Cell Res. 1997;232:25–28. - PubMed
-
- Shinohara T., Taniwaki M., Ishida Y., Kawaichi M., Honjo T. Structure and chromosomal localization of the human PD-1 gene (PDCD1) Genomics. 1994;23:704–706. - PubMed
-
- Vivier E., Daeron M. Immunoreceptor tyrosine-based inhibition motifs. Immunol. Today. 1997;18:286–291. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
