

The protein encoded by the proto-oncogene DEK changes the topology of chromatin and reduces the efficiency of DNA replication in a chromatin-specific manner
Affiliations
- PMID: 10837023
- PMCID: PMC316669
Item in Clipboard
The protein encoded by the proto-oncogene DEK changes the topology of chromatin and reduces the efficiency of DNA replication in a chromatin-specific manner
Genes Dev.
.
Abstract
The structure of chromatin regulates the genetic activity of the underlying DNA sequence. We report here that the protein encoded by the proto-oncogene DEK, which is involved in acute myelogenous leukemia, induces alterations of the superhelical density of DNA in chromatin. The change in topology is observed with chromatin but not with naked DNA and does not involve dissociation of core histones from chromatin. Moreover, these effects require histone H2A/H2B dimers in addition to histone H3/H4. We additionally tested whether the DEK protein affects DNA-utilizing processes and found that the DEK protein substantially reduces the replication efficiency of chromatin but not of naked DNA templates.
Figures
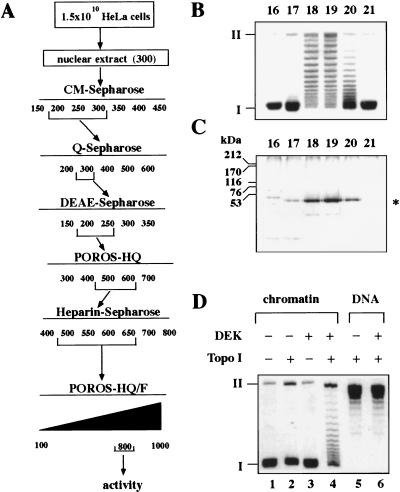
A 50-kD protein reduces the superhelical density of chromatin. (A) Purification scheme of the activity. Numbers indicate NaCl concentration (mm ). (B) SV40 chromatin was incubated with the fractions (16–21) from the POROS-HQ/F column in the presence of a cytosolic S100 replication extract. Purified DNA was investigated by agarose gel electrophoresis and ethidium bromide staining. (I) Supercoiled; (II) relaxed closed circular and nicked DNA. (C) Protein composition of the POROS-HQ/F-fractions. Proteins of the fractions (16–21) were separated by SDS-PAGE and visualized by silver staining. The position of size markers and the 50-kD protein (asterisk) is indicated. The faster-migrating minor band on the SDS-polyacrylamide gel most likely consists of a degradation product of the 50-kD protein, as its amount increased after prolonged incubation at 37°C. (D) Topoisomerase dependence of the reaction. SV40 chromatin (lanes 1–4) or relaxed SV40 DNA (lanes 5,6) were incubated in the absence or presence of topoisomerase I and purified DEK protein. Purified products were analyzed by agarose gel electrophoresis and visualized by ethidium bromide staining.
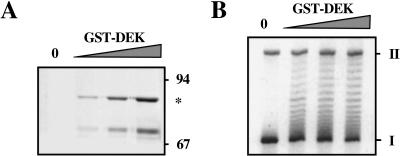
Recombinant GST–DEK changes the superhelical density of DNA in chromatin. (A) Increasing amounts of GST–DEK were separated by SDS-PAGE and visualized by silver staining. The positions of size markers and GST–DEK (*) are indicated. Lower band consists of a degradation product. (B) DEK assay with GST–DEK. SV40 chromatin was incubated with increasing amounts of GST–DEK in the presence of topoisomerase I. Purified DNA was investigated by agarose gel electrophoresis and ethidium bromide staining. (I) Supercoiled; (II) relaxed closed circular and nicked DNA.
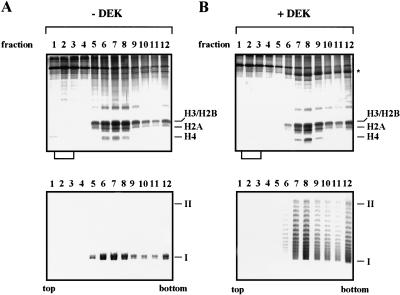
The DEK-induced change in linking number is not due to histone displacement. SV40 minichromosomes were incubated in the absence (A) or presence (B) of DEK and topoisomerase I and separated on glycerol gradients. Proteins of the gradient fractions (lanes 1–12) were analyzed by SDS-PAGE (top) and the DNA topology by agarose gel electrophoresis (bottom). The position of the core histones (H3, H2B, H2A, and H4) and supercoiled (I) and relaxed closed circular and nicked DNA (II) are indicated. Brackets below the SDS-PAGE show the position in which free histones are expected to appear. (Asterisk) Position of DEK protein.
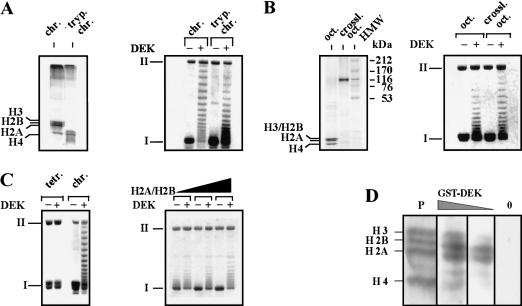
The DEK-induced change in topology depends on the presence of H2A/H2B dimers. (A, left) The amino-terminal histone domains of SV40 minichromosomes were removed by trypsin. Polypeptides were investigated by SDS-PAGE and visualized by silver staining. (Right) Non-treated chromatin (chr.) and trypsinized chromatin (tryp. chrom.) was used as substrate in the DEK assay. (B, left) Histone octamers (oct.) were cross-linked with DMS (crossl. oct.) and analyzed by SDS-PAGE stained with Coomassie brilliant blue. (HMW) High molecular weight marker in kilodaltons. (Right) SV40 DNA was reconstituted with control or cross-linked octamers and used as substrate in the DEK assay. (C, left) SV40 DNA was reconstituted with purified H3/H4 tetramers (tetr.) or histone octamers (chrom.). Chromatin was incubated in the absence or presence of DEK. (Right) H3/H4 containing chromatin was reconstituted with increasing amounts of H2A/H2B dimers and used as substrate in the DEK assay. (D) Far-Western blot analysis. Core histones were separated on an SDS–polyacrylamide gel, electroblotted, and renatured. The membrane was stained with Ponceau red (P) and incubated with decreasing amounts of GST–DEK (100 ng/cm2; 25 ng/cm2) and without DEK (0). The position of the core histones are indicated.
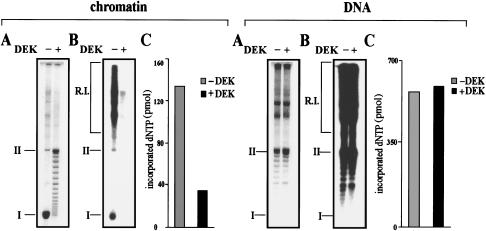
DEK inhibits the replication of chromatin but not of protein-free DNA. (A) SV40 DNA and SV40 chromatin were preincubated with DEK and topoisomerase I and used as templates in the SV40 in vitro replication system. Purified DNA was separated on an agarose gel and visualized by ethidium bromide staining. (B) Autoradiography of the agarose gel shown in A. The position of replicative intermediates (R.I.) of supercoiled (I) and relaxed closed circular and nicked molecules (II) are indicated. (C) Nucleotide incorporation (pmole) of the samples shown in B, as determined by TCA precipitation.
Similar articles
-
Daxx and histone deacetylase II associate with chromatin through an interaction with core histones and the chromatin-associated protein Dek.J Cell Sci. 2002 Aug 15;115(Pt 16):3319-30. doi: 10.1242/jcs.115.16.3319. J Cell Sci. 2002. PMID: 12140263
-
Bacterial Growth Inhibition Screen (BGIS) identifies a loss-of-function mutant of the DEK oncogene, indicating DNA modulating activities of DEK in chromatin.FEBS Lett. 2021 May;595(10):1438-1453. doi: 10.1002/1873-3468.14070. Epub 2021 Mar 24. FEBS Lett. 2021. PMID: 33686684
-
High-affinity interaction of poly(ADP-ribose) and the human DEK oncoprotein depends upon chain length.Biochemistry. 2010 Aug 24;49(33):7119-30. doi: 10.1021/bi1004365. Biochemistry. 2010. PMID: 20669926 Free PMC article.
-
Control of tumorigenesis and chemoresistance by the DEK oncogene.Clin Cancer Res. 2010 Jun 1;16(11):2932-8. doi: 10.1158/1078-0432.CCR-09-2330. Epub 2010 May 25. Clin Cancer Res. 2010. PMID: 20501624 Free PMC article. Review.
-
The DEK protein--an abundant and ubiquitous constituent of mammalian chromatin.Gene. 2004 Dec 8;343(1):1-9. doi: 10.1016/j.gene.2004.08.029. Gene. 2004. PMID: 15563827 Review.
Cited by
-
Intercellular trafficking of the nuclear oncoprotein DEK.Proc Natl Acad Sci U S A. 2013 Apr 23;110(17):6847-52. doi: 10.1073/pnas.1220751110. Epub 2013 Apr 8. Proc Natl Acad Sci U S A. 2013. PMID: 23569252 Free PMC article.
-
Doxorubicin induces prolonged DNA damage signal in cells overexpressing DEK isoform-2.PLoS One. 2022 Oct 3;17(10):e0275476. doi: 10.1371/journal.pone.0275476. eCollection 2022. PLoS One. 2022. PMID: 36190960 Free PMC article.
-
Overexpression of the cellular DEK protein promotes epithelial transformation in vitro and in vivo.Cancer Res. 2009 Mar 1;69(5):1792-9. doi: 10.1158/0008-5472.CAN-08-2304. Epub 2009 Feb 17. Cancer Res. 2009. PMID: 19223548 Free PMC article.
-
DEK associates with tumor stage and outcome in HPV16 positive oropharyngeal squamous cell carcinoma.Oncotarget. 2017 Apr 4;8(14):23414-23426. doi: 10.18632/oncotarget.15582. Oncotarget. 2017. PMID: 28423581 Free PMC article.
-
IRAK1 is a novel DEK transcriptional target and is essential for head and neck cancer cell survival.Oncotarget. 2015 Dec 22;6(41):43395-407. doi: 10.18632/oncotarget.6028. Oncotarget. 2015. PMID: 26527316 Free PMC article.
References
-
- Alexiadis V, Halmer L, Gruss C. Influence of core histone acetylation on SV40 minichromosomes replication in vitro. Chromosoma. 1997;105:324–331. - PubMed
-
- Ausio J, Dong F, van Holde KE. Use of selectively trysinized nucleosome core particles to analyze the role of the histone “tails” in the stabilization of the nucleosome. J Mol Biol. 1989;206:451–463. - PubMed
-
- Cairns BR, Lorch Y, Li Y, Zhang M, Lacomis L, Erdjument-Bromage H, Tempst P, Du J, Laurent B, Kornberg RD. RSC, an essential, abundant chromatin-remodeling complex. Cell. 1996;87:1249–1260. - PubMed
-
- Chang L, Loranger SS, Mizzen C, Ernst SG, Allis CD, Annunziato AT. Histones in transit: Cytosolic histone complexes and diacetylation of H4 during nucleosome assembly in human cells. Biochemistry. 1997;36:469–480. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous