

Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion
- PMID: 10805752
- PMCID: PMC85780
- DOI: 10.1128/MCB.20.11.4106-4114.2000
Analysis of fractalkine receptor CX(3)CR1 function by targeted deletion and green fluorescent protein reporter gene insertion
Abstract
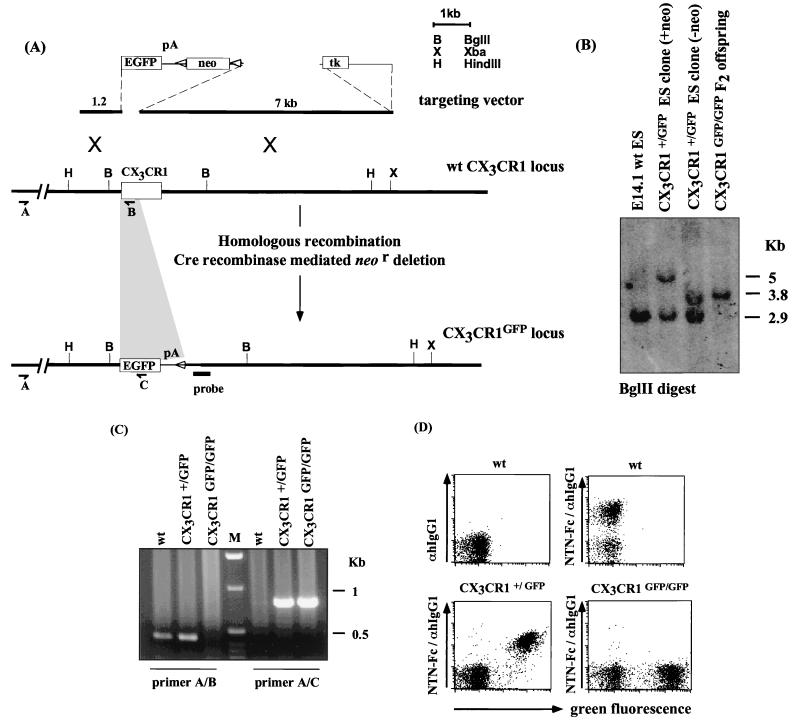
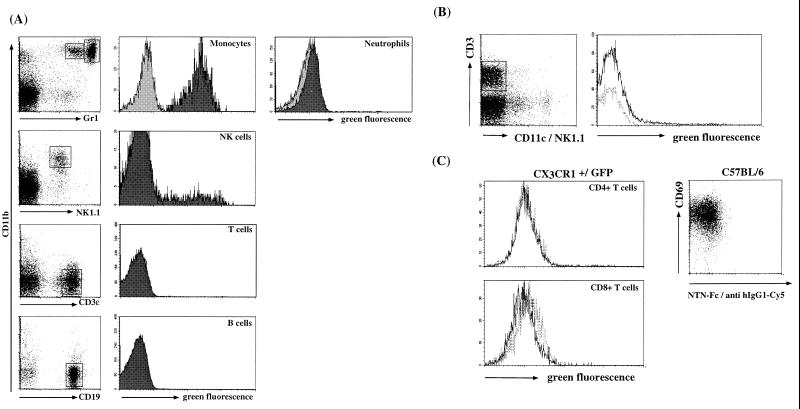
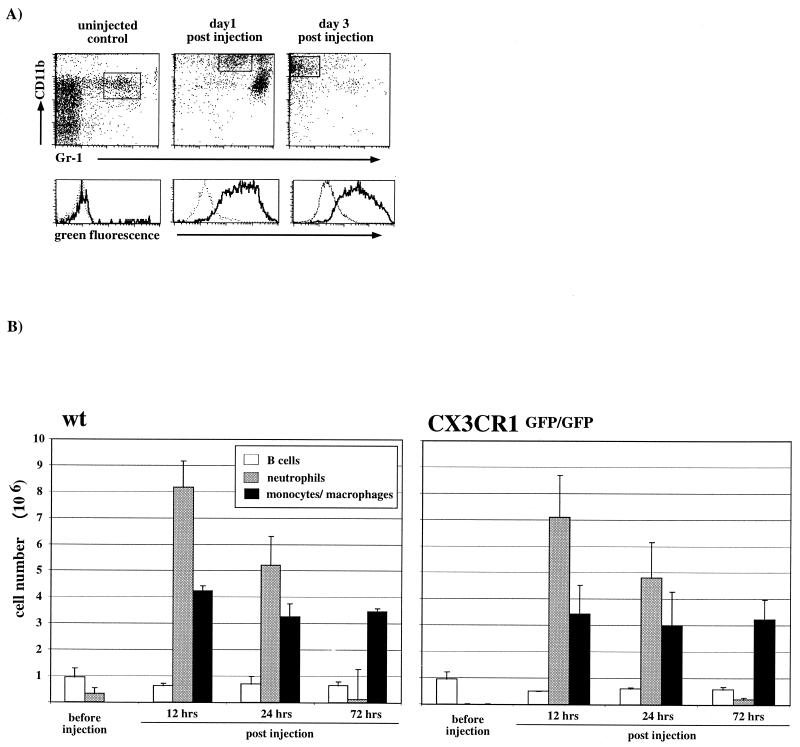
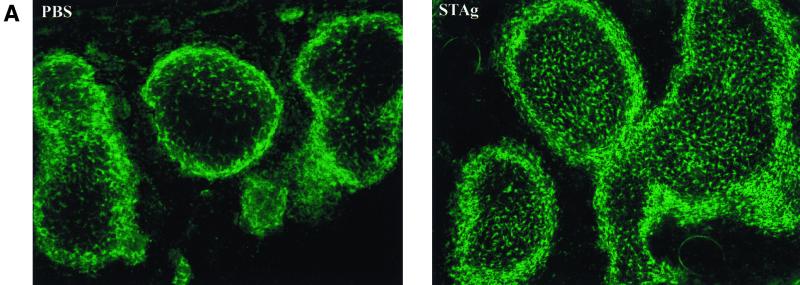
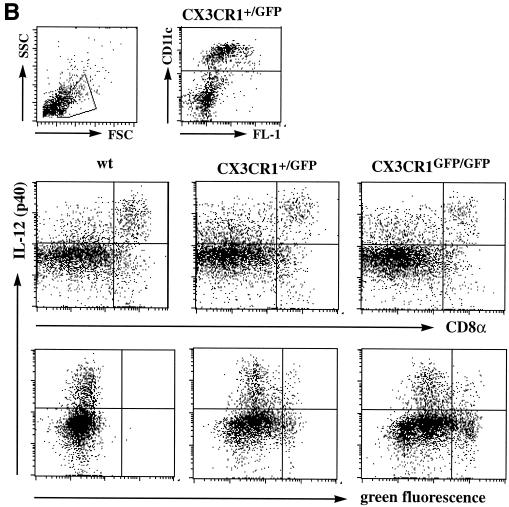
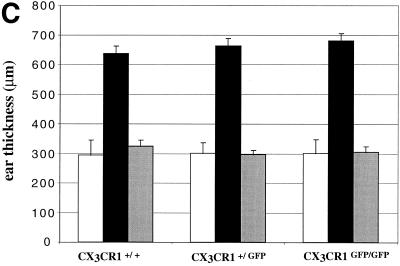
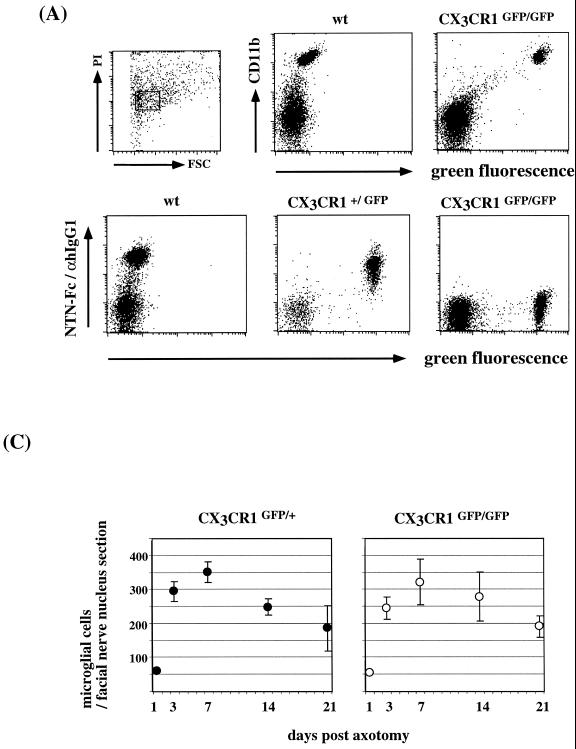
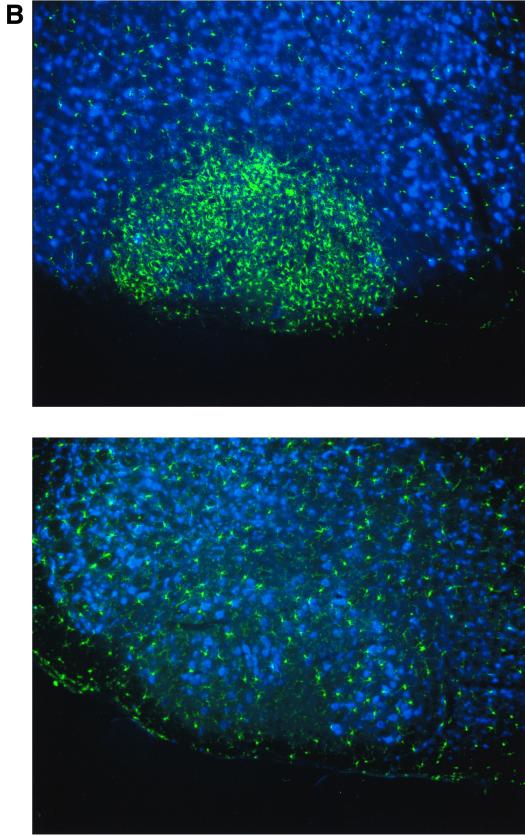
The seven-transmembrane receptor CX(3)CR1 is a specific receptor for the novel CX(3)C chemokine fractalkine (FKN) (neurotactin). In vitro data suggest that membrane anchoring of FKN, and the existence of a shed, soluble FKN isoform allow for both adhesive and chemoattractive properties. Expression on activated endothelium and neurons defines FKN as a potential target for therapeutic intervention in inflammatory conditions, particularly central nervous system diseases. To investigate the physiological function of CX(3)CR1-FKN interactions, we generated a mouse strain in which the CX(3)CR1 gene was replaced by a green fluorescent protein (GFP) reporter gene. In addition to the creation of a mutant CX(3)CR1 locus, this approach enabled us to assign murine CX(3)CR1 expression to monocytes, subsets of NK and dendritic cells, and the brain microglia. Analysis of CX(3)CR1-deficient mice indicates that CX(3)CR1 is the only murine FKN receptor. Yet, defying anticipated FKN functions, absence of CX(3)CR1 interferes neither with monocyte extravasation in a peritonitis model nor with DC migration and differentiation in response to microbial antigens or contact sensitizers. Furthermore, a prominent response of CX(3)CR1-deficient microglia to peripheral nerve injury indicates unimpaired neuronal-glial cross talk in the absence of CX(3)CR1.
Figures
Similar articles
-
Neuron-glia crosstalk in health and disease: fractalkine and CX3CR1 take centre stage.Open Biol. 2013 Dec 18;3(12):130181. doi: 10.1098/rsob.130181. Open Biol. 2013. PMID: 24352739 Free PMC article. Review.
-
Expression of fractalkine and its receptor, CX3CR1, in atopic dermatitis: possible contribution to skin inflammation.J Allergy Clin Immunol. 2004 May;113(5):940-8. doi: 10.1016/j.jaci.2004.02.030. J Allergy Clin Immunol. 2004. PMID: 15131578
-
Contribution of platelet CX(3)CR1 to platelet-monocyte complex formation and vascular recruitment during hyperlipidemia.Arterioscler Thromb Vasc Biol. 2012 May;32(5):1186-93. doi: 10.1161/ATVBAHA.111.243485. Epub 2012 Mar 1. Arterioscler Thromb Vasc Biol. 2012. PMID: 22383701
-
Molecular cloning and characterization of canine fractalkine and its receptor CX3CR1.Vet Immunol Immunopathol. 2012 Jan 15;145(1-2):100-9. doi: 10.1016/j.vetimm.2011.10.018. Epub 2011 Nov 17. Vet Immunol Immunopathol. 2012. PMID: 22169328
-
Fractalkine--a local inflammatory marker aggravating platelet activation at the vulnerable plaque.Thromb Haemost. 2012 Sep;108(3):457-63. doi: 10.1160/TH12-04-0271. Epub 2012 Jun 28. Thromb Haemost. 2012. PMID: 22739755 Review.
Cited by
-
Autophagy protein ULK1 interacts with and regulates SARM1 during axonal injury.Proc Natl Acad Sci U S A. 2022 Nov 22;119(47):e2203824119. doi: 10.1073/pnas.2203824119. Epub 2022 Nov 14. Proc Natl Acad Sci U S A. 2022. PMID: 36375051 Free PMC article.
-
Characterisation of C101248: A novel selective THIK-1 channel inhibitor for the modulation of microglial NLRP3-inflammasome.Neuropharmacology. 2023 Feb 15;224:109330. doi: 10.1016/j.neuropharm.2022.109330. Epub 2022 Nov 12. Neuropharmacology. 2023. PMID: 36375694 Free PMC article.
-
Time-lapse Whole-field Fluorescence Imaging of Microglia ProcessesMotility in Acute Mouse Hippocampal Slices and Analysis.Bio Protoc. 2019 Apr 20;9(8):e3220. doi: 10.21769/BioProtoc.3220. eCollection 2019 Apr 20. Bio Protoc. 2019. PMID: 33655009 Free PMC article.
-
CX3 chemokine receptor 1 deficiency leads to reduced dendritic complexity and delayed maturation of newborn neurons in the adult mouse hippocampus.Neural Regen Res. 2015 May;10(5):772-7. doi: 10.4103/1673-5374.156979. Neural Regen Res. 2015. PMID: 26109952 Free PMC article.
-
High fat diet causes depletion of intestinal eosinophils associated with intestinal permeability.PLoS One. 2015 Apr 2;10(4):e0122195. doi: 10.1371/journal.pone.0122195. eCollection 2015. PLoS One. 2015. PMID: 25837594 Free PMC article.
References
-
- Bazan J F, Bacon K B, Hardiman G, Wang W, Soo K, Rossi D, Greaves D R, Zlotnik A, Schall T J. A new class of membrane-bound chemokine with a CX3C motif. Nature. 1997;385:640–644. - PubMed
-
- Butcher E C, Picker L J. Lymphocyte homing and homeostasis. Science. 1996;272:60–66. - PubMed
-
- Campbell J J, Hedrick J, Zlotnik A, Siani M A, Thompson D A, Butcher E C. Chemokines and the arrest of lymphocytes rolling under flow conditions. Science. 1998;279:381–384. - PubMed
-
- Foussat A, Coulomb-L'Hermine A, Gosling J, Krzysiek R, Durand-Gasselin I, Schall T, Balian A, Richard Y, Galanaud P, Emilie D. Fractalkine receptor expression by T lymphocyte subpopulations and in vivo production of fractalkine in human. Eur J Immunol. 2000;30:87–97. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous