

Attenuation of apoptotic cell detection triggers thymic regeneration after damage
- PMID: 34610317
- PMCID: PMC8627669
- DOI: 10.1016/j.celrep.2021.109789
Attenuation of apoptotic cell detection triggers thymic regeneration after damage
Abstract
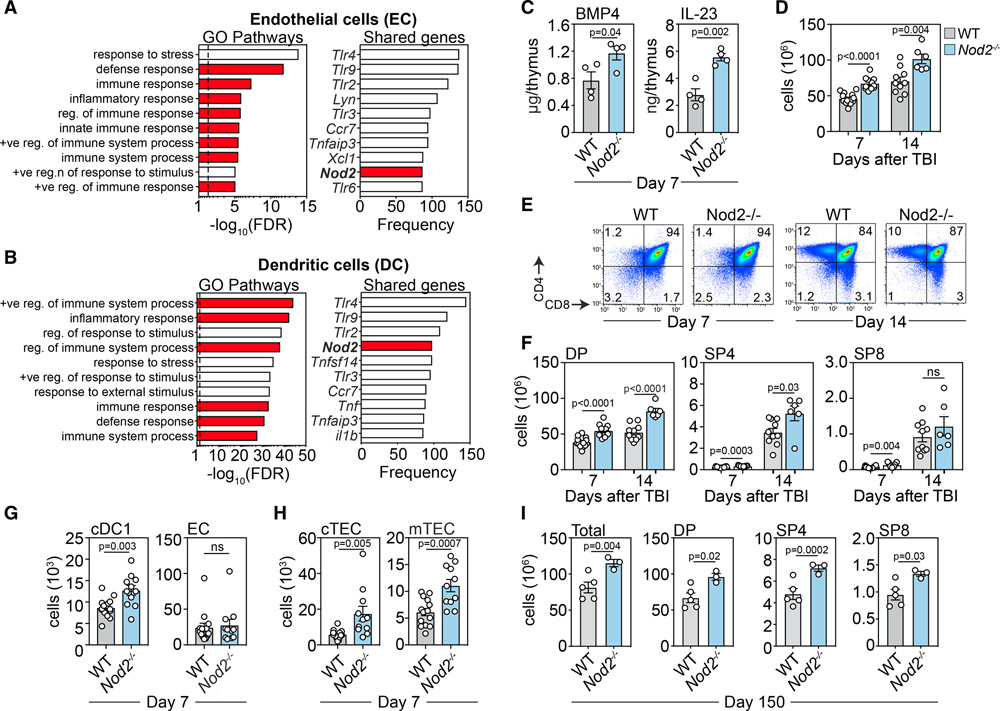
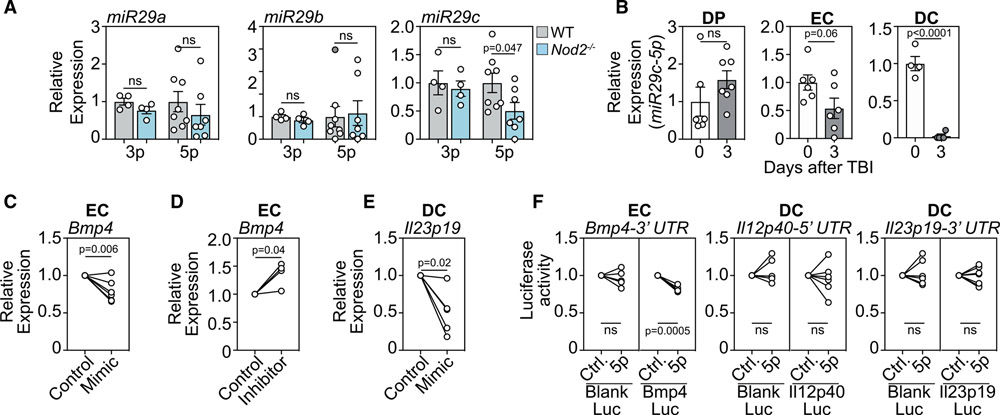
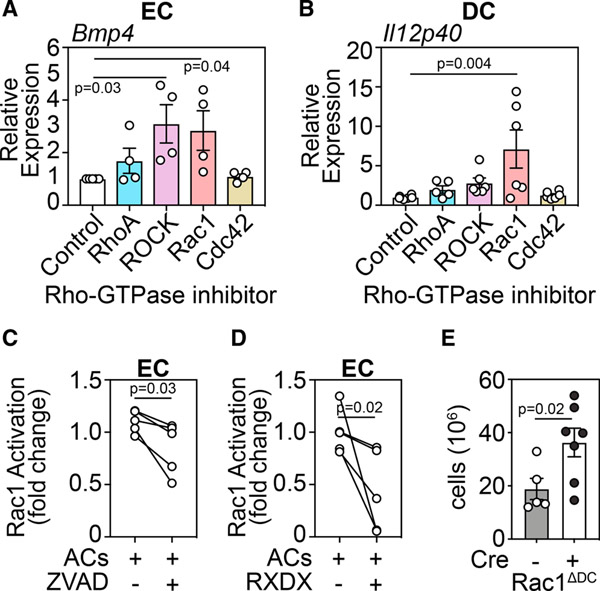
The thymus, which is the primary site of T cell development, is particularly sensitive to insult but also has a remarkable capacity for repair. However, the mechanisms orchestrating regeneration are poorly understood, and delayed repair is common after cytoreductive therapies. Here, we demonstrate a trigger of thymic regeneration, centered on detecting the loss of dying thymocytes that are abundant during steady-state T cell development. Specifically, apoptotic thymocytes suppressed production of the regenerative factors IL-23 and BMP4 via TAM receptor signaling and activation of the Rho-GTPase Rac1, the intracellular pattern recognition receptor NOD2, and micro-RNA-29c. However, after damage, when profound thymocyte depletion occurs, this TAM-Rac1-NOD2-miR29c pathway is attenuated, increasing production of IL-23 and BMP4. Notably, pharmacological inhibition of Rac1-GTPase enhanced thymic function after acute damage. These findings identify a complex trigger of tissue regeneration and offer a regenerative strategy for restoring immune competence in patients whose thymic function has been compromised.
Keywords: NOD2; Rac1 GTPase; T cell development; TAM receptors; apoptotic cell death; lymphopenia; thymus; tissue regeneration.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.K. and J.A.D. have filed a patent application on this work.
Figures


Similar articles
-
Production of BMP4 by endothelial cells is crucial for endogenous thymic regeneration.Sci Immunol. 2018 Jan 12;3(19):eaal2736. doi: 10.1126/sciimmunol.aal2736. Sci Immunol. 2018. PMID: 29330161 Free PMC article.
-
Interleukin-22 drives endogenous thymic regeneration in mice.Science. 2012 Apr 6;336(6077):91-5. doi: 10.1126/science.1218004. Epub 2012 Mar 1. Science. 2012. PMID: 22383805 Free PMC article.
-
When the Damage Is Done: Injury and Repair in Thymus Function.Front Immunol. 2020 Aug 12;11:1745. doi: 10.3389/fimmu.2020.01745. eCollection 2020. Front Immunol. 2020. PMID: 32903477 Free PMC article. Review.
-
Regenerative capacity of adult cortical thymic epithelial cells.Proc Natl Acad Sci U S A. 2012 Feb 28;109(9):3463-8. doi: 10.1073/pnas.1118823109. Epub 2012 Feb 13. Proc Natl Acad Sci U S A. 2012. PMID: 22331880 Free PMC article.
-
Repairing thymic function.Curr Opin Organ Transplant. 2013 Jun;18(3):363-8. doi: 10.1097/MOT.0b013e3283615df9. Curr Opin Organ Transplant. 2013. PMID: 23660780 Review.
Cited by
-
The thymus road to a T cell: migration, selection, and atrophy.Front Immunol. 2024 Aug 27;15:1443910. doi: 10.3389/fimmu.2024.1443910. eCollection 2024. Front Immunol. 2024. PMID: 39257583 Free PMC article. Review.
-
Immune tolerance and the prevention of autoimmune diseases essentially depend on thymic tissue homeostasis.Front Immunol. 2024 Mar 20;15:1339714. doi: 10.3389/fimmu.2024.1339714. eCollection 2024. Front Immunol. 2024. PMID: 38571951 Free PMC article. Review.
-
Disorganization of secondary lymphoid organs and dyscoordination of chemokine secretion as key contributors to immune aging.Semin Immunol. 2023 Nov;70:101835. doi: 10.1016/j.smim.2023.101835. Epub 2023 Aug 29. Semin Immunol. 2023. PMID: 37651849 Free PMC article. Review.
-
Activation of the zinc-sensing receptor GPR39 promotes T-cell reconstitution after hematopoietic cell transplant in mice.Blood. 2022 Jun 23;139(25):3655-3666. doi: 10.1182/blood.2021013950. Blood. 2022. PMID: 35357432 Free PMC article.
-
Eosinophils are an essential element of a type 2 immune axis that controls thymus regeneration.Sci Immunol. 2022 Mar 11;7(69):eabn3286. doi: 10.1126/sciimmunol.abn3286. Epub 2022 Mar 11. Sci Immunol. 2022. PMID: 35275754 Free PMC article.
References
-
- Abramson J, and Anderson G (2017). Thymic Epithelial Cells. Annu. Rev. Immunol 35, 85–118. - PubMed
-
- Bartel DP (2004). MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116, 281–297. - PubMed
-
- Billmann-Born S, Till A, Arlt A, Lipinski S, Sina C, Latiano A, Annese V, Häsler R, Kerick M, Manke T, et al. (2011). Genome-wide expression profiling identifies an impairment of negative feedback signals in the Crohn’s disease-associated NOD2 variant L1007fsinsC. J. Immunol 186, 4027–4038. - PubMed
-
- Bosch M, Khan FM, and Storek J (2012). Immune reconstitution after hematopoietic cell transplantation. Curr. Opin. Hematol 19, 324–335. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
